Dr J.P. Schadé - De functie van het zenuwstelsel
Aula Boeken, Het Spectrum, 1968Over het boek: Dit helder geschreven en rijk geïllustreerde boek biedt een overzicht van het zo boeiende en zich buitengewoon snel ontwikkelende wetenschapsgebied van de moleculaire neuro-biologie, voornamelijk met betrekking tot de functie en structuur van het zenuwstelsel bij mens en dier. Hoewel het boek in de eerste plaats bedoeld is als een inleiding tot de moderne neurofysiologie voor studenten en afgestudeerden in de geneeskunde, biologie, andere natuurwetenschappen en psychologie, geeft het ook een wetenschappelijk verantwoorde handleiding waarmee de ontwikkelde lezer zich een inzicht kan verwerven in datgene, wat een waardevol en integrerend deel van heel zijn bestaan uitmaakt: de werking van het zenuwstelsel en de zintuigen.

Dr. J.P. Schadé
Inhoud
Voorwoord
1. Ultrastructuur 9
Ultratechnieken
Het neuron - Structuur
Nucleïnezuren in het neuron
Endoplasmatisch reticulum en mitochondriën
Organisatie van het neuronmembraan
Moleculaire membraanstructuur
Schakelplaats in het zenuwstelsel: de synaps
Unieke betekenis van neurotransmitters
2. Elektrofysiologie 27
Zenuwstelsel: informatieverwerkende rekenmachine
De reflexkring
Informatie-overdracht in een reflexkring
De reflex: een onwillekeurige reactie
De receptoren-zintuigcellen
Potentiaalverschil in de zintuigcel
Ionenverschuivingen in de zintuigcel
De actiepotentiaal 1
De actiepotentiaal 2
Generatorpotentialen
Geleidingssnelheid
De rustpotentiaal van de zenuwcel
Codering in een axon
Antagonisten en synergisten
De motorische zenuwcel
Ionen en rustpotentiaal van de synaps
Prikkel van receptor naar synaps: excitatie en inhibitie
Excitatie door een transmitterstof
Exciterende potentialen
Inhibitie door een transmitterstof
Inhiberende potentialen
De zenuwcel: een microscopische rekenmachine
Prikkeloverdracht van zenuw op spier
Elektronenmicroscopie van de spiervezel
De neuromusculaire synaps
Spierpotentialen
Van spierpotentiaal tot spiercontractie
1. Ultrastructuur 9
Ultratechnieken
Het neuron - Structuur
Nucleïnezuren in het neuron
Endoplasmatisch reticulum en mitochondriën
Organisatie van het neuronmembraan
Moleculaire membraanstructuur
Schakelplaats in het zenuwstelsel: de synaps
Unieke betekenis van neurotransmitters
2. Elektrofysiologie 27
Zenuwstelsel: informatieverwerkende rekenmachine
De reflexkring
Informatie-overdracht in een reflexkring
De reflex: een onwillekeurige reactie
De receptoren-zintuigcellen
Potentiaalverschil in de zintuigcel
Ionenverschuivingen in de zintuigcel
De actiepotentiaal 1
De actiepotentiaal 2
Generatorpotentialen
Geleidingssnelheid
De rustpotentiaal van de zenuwcel
Codering in een axon
Antagonisten en synergisten
De motorische zenuwcel
Ionen en rustpotentiaal van de synaps
Prikkel van receptor naar synaps: excitatie en inhibitie
Excitatie door een transmitterstof
Exciterende potentialen
Inhibitie door een transmitterstof
Inhiberende potentialen
De zenuwcel: een microscopische rekenmachine
Prikkeloverdracht van zenuw op spier
Elektronenmicroscopie van de spiervezel
De neuromusculaire synaps
Spierpotentialen
Van spierpotentiaal tot spiercontractie
3. Neuronale circuits 85
Neuronale plasticiteit
Functie van inhibitie in neuronale integratie
De sturende taak van antidrome inhibitie
Presynaptische inhibitie in een axo-axonale synaps
Dubbele inhibitie veroorzaakt facilitatie
Schakelmogelijkheden voor 15 miljoen eenheden
4. Neorotransmitters 95
De chemische schakel in informatie-overdracht
Analoog-digitaal-analoog-digitaal transmissie
Het autonome zenuwstelsel
Cholinerge en adrenerge neuronen
Biogene aminen als neurotransmitters
Catecholaminen
Fenylalanine → tyrosine
Tyrosine → DOPA
DOPA → dopamine
Dopamine en de perifere circulatie
Dopamine → noradrenaline → adrenaline
De afbraak van catecholaminen
Monoamine-oxydasen (MAO)
MAO remmers
Overzicht van de werking van de catecholaminen
Catecholaminendepots
Opname van catecholaminen in het neuron
Opname van catecholaminen in de granulae
De afgifte van noradrenaline
MAO en MAO-remmers
Reserpine-effect
De organisatie van het autonome zenuwstelsel
5. Lijst van termen 157
Neuronale plasticiteit
Functie van inhibitie in neuronale integratie
De sturende taak van antidrome inhibitie
Presynaptische inhibitie in een axo-axonale synaps
Dubbele inhibitie veroorzaakt facilitatie
Schakelmogelijkheden voor 15 miljoen eenheden
4. Neorotransmitters 95
De chemische schakel in informatie-overdracht
Analoog-digitaal-analoog-digitaal transmissie
Het autonome zenuwstelsel
Cholinerge en adrenerge neuronen
Biogene aminen als neurotransmitters
Catecholaminen
Fenylalanine → tyrosine
Tyrosine → DOPA
DOPA → dopamine
Dopamine en de perifere circulatie
Dopamine → noradrenaline → adrenaline
De afbraak van catecholaminen
Monoamine-oxydasen (MAO)
MAO remmers
Overzicht van de werking van de catecholaminen
Catecholaminendepots
Opname van catecholaminen in het neuron
Opname van catecholaminen in de granulae
De afgifte van noradrenaline
MAO en MAO-remmers
Reserpine-effect
De organisatie van het autonome zenuwstelsel
5. Lijst van termen 157
Voorwoord
In dit boek, dat als een inleiding tot de kennis van de functie van het zenuwstelsel beschouwd moet worden, zal de nadruk worden gelegd op de elektrofysiologie van de neuronen.
Een van de boeiendste onderwerpen uit de hedendaagse neurobiologie is de communicatie tussen zenuwcellen en het transport van informatie in het perifere en centrale zenuwstelsel. Deze processen spelen zich af op moleculair niveau en kunnen met de thans ter beschikking staande apparatuur, zoals elektronenmicroscoop en rekenmachine, tot in de kleinste details worden geanalyseerd. Het is dan ook niet ten onrechte dat we thans spreken van een nieuwe wetenschap: de moleculaire neurobiologie.
Bij het samenstellen van dit boek is geen enkele poging gedaan om een samenvattend geheel te geven, hiervoor zou zelfs een omvang die duizendmaal zo groot is als de huidige, nog onvoldoende zijn. Getracht is alleen een inleiding te geven in de moderne neurofysiologie, tegen de achtergrond van de structuur en de biochemische eigenschappen van het zenuwstelsel. Voor een volledige bestudering van de behandelde onderwerpen wordt de lezer verwezen naar de bibliografie.
Bijzondere dank wordt gebracht aan C.J. van der Grond voor de vele tekeningen van de eerste drie hoofdstukken en aan mej. J. Sels voor het vele typewerk.
J.P. Schadé
terug naar de Inhoud
1. Ultrastructuur
De tien tot vijftien miljard zennuwcellen van de hersenen en het ruggemerg vormen tezamen een ingewikkeld conglomerant van grijze en witte stof, opgeborgen in de schedelruimte en het wervelkanaal.
Het elektronenmicroscoop heeft een geheel nieuwe wereld ontvouwd in het binnenste van de zenuwcellen. De resultaten van dit onderzoek zijn van grote waarde voor de interpretatie van de fysiologische en biochemische benadering van het zenuwstelsel. Elke zenuwcel is een structurele en functionele eenheid. Het ultramicroscopische onderzoek heeft een tip van de sluier opgelicht, waardoor een complexe structuur van membranen en kanalen onthuld werd, die het morfologische substraat vormen van een ingewikkeld systeem van functies zoals eiwitsynthese, productie van neurotransmitters, aanmaak van enzymen, enz.
Dit hoofdstuk geeft een inleiding in enkele bouwprincipes van het neuron, waarbij de nadruk valt op de ultramicroscopische bouw, zoals die met behulp van het elektronenmicroscoop onderzocht wordt.
Ultratechnieken
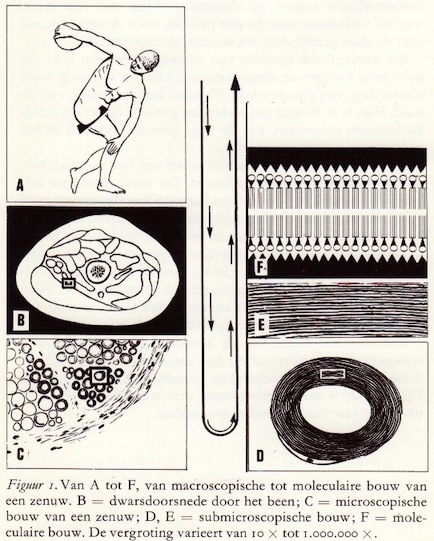 Zowel het fundamentele als het klinische onderzoek van het zenuwstelsel heeft in de laatste twee decennia een stormachtige ontwikkeling doorgemaakt. De klinische - over het algemeen instrumentloze - diagnostiek is steeds meer gebruik gaan maken van de mogelijkheden die de moderne technologie biedt. Zo hebben sinds de tweede wereldoorlog een aantal technieken voor diagnostiek en therapie hun intrede gedaan. Wij noemen hiervan de elektromyografie (het elektrisch onderzoek van de spieren), de stereotactische ingrepen zoals de cryochirurgische behandeling van de ziekte van Parkinson en het gebruik van radioactieve isotopen. De introductie van deze methoden heeft een accentverschuiving teweeg gebracht naar de meer mechanische of instrumentele benadering van de diagnostiek en behandeling van ziekten van het zenuwstelsel, zonder echter de zuiver klinische diagnostiek in waarde te doen verminderen. Ook van geheel andere zijde worden belangrijke resultaten geboekt, daar zowel de neurochemie als de neurofarmacologie langzaam maar zeker de kinderschoenen ontgroeien.
Zowel het fundamentele als het klinische onderzoek van het zenuwstelsel heeft in de laatste twee decennia een stormachtige ontwikkeling doorgemaakt. De klinische - over het algemeen instrumentloze - diagnostiek is steeds meer gebruik gaan maken van de mogelijkheden die de moderne technologie biedt. Zo hebben sinds de tweede wereldoorlog een aantal technieken voor diagnostiek en therapie hun intrede gedaan. Wij noemen hiervan de elektromyografie (het elektrisch onderzoek van de spieren), de stereotactische ingrepen zoals de cryochirurgische behandeling van de ziekte van Parkinson en het gebruik van radioactieve isotopen. De introductie van deze methoden heeft een accentverschuiving teweeg gebracht naar de meer mechanische of instrumentele benadering van de diagnostiek en behandeling van ziekten van het zenuwstelsel, zonder echter de zuiver klinische diagnostiek in waarde te doen verminderen. Ook van geheel andere zijde worden belangrijke resultaten geboekt, daar zowel de neurochemie als de neurofarmacologie langzaam maar zeker de kinderschoenen ontgroeien.Wat het fundamentele onderzoek van het zenuwstelsel betreft, is er niet zo zeer sprake van een accentverschuiving als wel van een dimensieverschuiving. De verschillende disciplines zoals de neurofysiologie, neurocybernetica, neuroanatomie, neuro-endocrinologie etc., die onder de algemene noemer 'neurobiologie' tezamen de basiswetenschappen van de neurologie zijn, hebben in zeer grote mate gebruik gemaakt van de toepassing van elektronische meetapparatuur.
Was er een tiental jaren terug nog slechts sprake van een macro- en een microneurobiologie, thans hebben de elektronenmicroscoop, de ultracentrifuge en de elektronische rekenmachine, om maar een aantal voorbeelden te noemen, ons in staat gesteld om door te dringen tot het moleculaire niveau waarop de levensprocessen zich afspelen. We kunnen dan ook terecht spreken van een nieuwe wetenschap: de moleculaire neurobiologie. (10)
Het neuron - Structuur
 De structurele en functionele eenheid van het zenuwstelsel is de zenuwcel. Cellen van andere organen zoals de nieren en de lever hebben een relatief eenvoudige vorm in vergelijking met de zenuwcel. Het meest karakteristieke verschil is wel het grote aantal uitlopers, die verschillend van lengte kunnen zijn.
De structurele en functionele eenheid van het zenuwstelsel is de zenuwcel. Cellen van andere organen zoals de nieren en de lever hebben een relatief eenvoudige vorm in vergelijking met de zenuwcel. Het meest karakteristieke verschil is wel het grote aantal uitlopers, die verschillend van lengte kunnen zijn.Wanneer wij spreken over het innerveren (prikkelen) van een spier door een zenuw bedoelen we het feit, dat de zenuwvezels van de desbetreffende zenuw op dusdanige wijze met het spierweefsel verbonden zijn dat een prikkel, door de zenuwvezels naar de spier geleid, deze tot samentrekking brengt.
We kunnen ook spreken van innervatie van een klier. In zo'n geval brengen de zenuwvezels de klier tot vorming en/of afscheiding van zijn product. Evenzo van innervatie van de huid. Hier is de functie net andersom gericht: de zenuwvezels brengen nu informatie van de huid naar de hersenen of het ruggemerg.
De zenuwcellen of neuronen zijn zowel wat hun vorm als hun functie betreft zeer gespecialiseerd. Dit heeft geleid tot cellen die bij uitstek in staat zijn tot het geleiden van impulsen of prikkels. Impulsen zijn bepaalde toestandsveranderingen van het neuron, die in het lichaam de betekenis hebben van boodschappen. De zenuwcel is hiervoor uitgerust met twee soorten uitlopers:
- de dendrieten, kleine, meestal rijkvertakte uitlopertjes die de impulsen opvangen,
- en de neuriet of het axon, die de impuls vervoert naar andere zenuwcellen of naar een uitvoerend orgaan zoals een spier of een klier.
Het klassieke beeld van de zenuwcel als zijnde een van uitlopers voorziene structuur met een kern en cytoplasma is door de diverse ultratechnieken grondig omgevormd. Elk neuron is een microcosmos, een wereldje op zichzelf, voorzien van tienduizenden organellen. (12)
Nucleïnezuren in het neuron
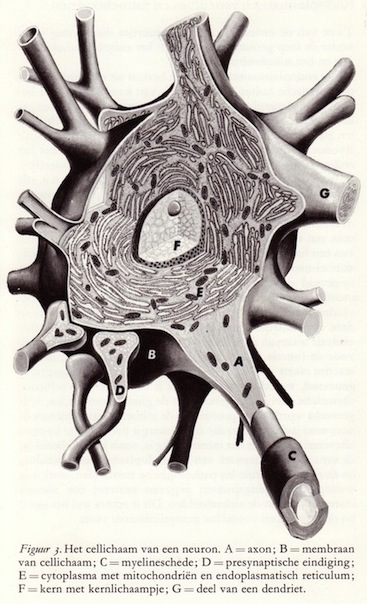 Het zenuwstelsel is opgebouwd uit vele biljoenen zenuwcellen, waarvan er geen twee gelijk zijn. Zoals bij elke dierlijke cel kunnen we bij oppervlakkige beschouwing een celwand en een celinhoud onderscheiden, waarvan de laatste een kern en de zgn. Nissl-lichaampjes bevat. Deze kern, niet alleen de drager van de erfelijke eigenschappen, speelt gedurende het celbestaan een onmisbare en leidende rol, zonder welke de cel niet in staat is zijn activiteiten voort te zetten. Daartoe bevat de kern een hoge concentratie nucleïnezuren, en wel de uiterst ingenieus opgebouwde desoxyribonucleïnezuren (afgekort DNZ of DNA, van desoxyribonucleic acid), waarin alle informatie voor de individu-specitieke eiwitten is vastgelegd en welke een essentiëel bestanddeel vormen van de chromosomen.
Het zenuwstelsel is opgebouwd uit vele biljoenen zenuwcellen, waarvan er geen twee gelijk zijn. Zoals bij elke dierlijke cel kunnen we bij oppervlakkige beschouwing een celwand en een celinhoud onderscheiden, waarvan de laatste een kern en de zgn. Nissl-lichaampjes bevat. Deze kern, niet alleen de drager van de erfelijke eigenschappen, speelt gedurende het celbestaan een onmisbare en leidende rol, zonder welke de cel niet in staat is zijn activiteiten voort te zetten. Daartoe bevat de kern een hoge concentratie nucleïnezuren, en wel de uiterst ingenieus opgebouwde desoxyribonucleïnezuren (afgekort DNZ of DNA, van desoxyribonucleic acid), waarin alle informatie voor de individu-specitieke eiwitten is vastgelegd en welke een essentiëel bestanddeel vormen van de chromosomen.Het compartiment van de cel buiten de kern, het cytoplasma, bevat eveneens een hoge concentratie van een nauwverwant nucleïnezuur: het ribonucleïnezuur (RNZ of RNA van ribonucleic acid). Grote agglomeraten van moleculen RNA zijn reeds lang geleden zichtbaar gemaakt en werden naar de ontdekker 'Nissl-lichaampjes' (ribosomen) genoemd. Het DNA en het RNA besturen met elkaar de functies van de cel en zijn tevens verantwoordelijk voor de productie van eiwitten, waardoor de cel tot op hoge leeftijd zijn structurele integriteit weet te handhaven. Vandaar dat deze stoffen terecht de levensmoleculen worden genoemd en het is dan ook niet verwonderlijk dat ze in hoge concentratie in de zenuwcellen voorkomen.
Van de vele andere organellen in het cytoplasma moeten hier ook nog worden genoemd het endoplasmarisch reticulum en de mitochondria (zie blz. 16), het Golgi-apparaat en vele kleine structuurtjes zoals pigment en vetkorretjes. Het Golgi-apparaat dient onder andere als voorraadschuur van materiaal voor de celmembranen; deze moeten voortdurend vernieuwd en gerepareerd worden en de bouwstenen hiervoor worden gefabriceerd in het Golgi-apparaat. (14)
Endoplasmatisch reticulum en mitochondriën
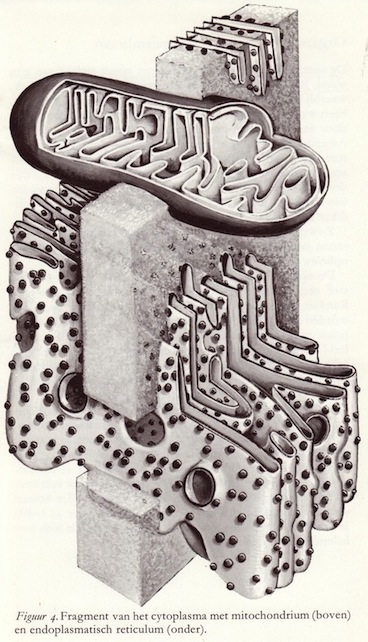 Twee van de neuronale ultrastructuurtjes dienen nog extra onder de loep te worden genomen: het endoplasmatisch reticulum en het mitochondrium.
Twee van de neuronale ultrastructuurtjes dienen nog extra onder de loep te worden genomen: het endoplasmatisch reticulum en het mitochondrium.Het endoplasmatisch reticulum bestaat uit een doolhof van concentrische holten. Dit netwerk van kanaaltjes wordt begrensd door membranen, waarin zich tegen de naar buiten gekeerde kant korrels bevinden. De naar binnen gekeerde kanten zijn glad. Elk van deze korrels is samengesteld uit ribonucleïnezuren en eiwitten. Ultracentrifugering doet het endoplasmatisch reticulum uiteenvallen in kleinere deeltjes, de z.g. microsomen. In deze microsomen worden o.a. de eiwitten gemaakt die niet alleen van belang zijn voor de structurele integriteit van de zenuwcellen maar ook voor de overdracht van informatie.
De mitochondriën hebben eveneens een specifiek uiterlijk. Het zijn langgerekte organellen van een halve tot enkele μ's lang, die omgeven zijn door een dubbel-membraan. Hiervan uitgaande wordt het binnenste opgevuld door ook een ingewikkeld stelsel van dubbelmembranen die men christae noemt.
In een grote zenuwcel bevinden zich 5.000 tot 10.000 van deze mitochondriën. Elk van deze structuurtjes kan vergeleken worden met een krachtcentrale, waarvan de belangrijkste taak is de energie te leveren voor het functioneren van de cel. In de mitochondriën vinden de reacties plaats van de Krebs-cyclus, ook wel citroenzuurcyclus genoemd, waardoor het mogelijk is dat grote hoeveelheden chemische energie, opgehoopt in de glucose-moleculen, vrijgemaakt worden. De positie van de mitochondriën binnen de zenuwcel is dan ook zo dat zij de energie precies daar kunnen afleveren waar deze het meest nodig is, zoals bijvoorbeeld bij de synapsen. Onderzoek van het endoplasmatisch reticulum en de mitochondriën bij pathologische toestanden heeft nog weinig aanknopingspunten gegeven omtrent een nieuwe interpretatie van de ziektebeelden. Dit is echter wel het geval bij de celwand en vooral het gemyeliniseerde axon. (16)
Organisatie van het neuronmembraan
 De specifieke functie van neuron speelt zich af aan zijn wand, het membraan. Elektronenmicroscopisch onderzoek, waarbij gebruik werd gemaakt van zeer hoge vergrotingen, heeft waarschijnlijk gemaakt dat elke dierlijke cel is omgeven door een membraan met identieke structuur. In de Anglo-Amerikaanse literatuur spreekt men van 'unit-membranestructure'. Op doorsnede kan men in de elektronenmicroscoop aan zo'n membraan drie lagen onderscheiden: twee donkere lagen ter dikte van ongeveer 20 Å, ter weerszijden van een lichte die ongeveer 55 Å dik is. De totale membraandikte bedraagt dus 75 Å.
De specifieke functie van neuron speelt zich af aan zijn wand, het membraan. Elektronenmicroscopisch onderzoek, waarbij gebruik werd gemaakt van zeer hoge vergrotingen, heeft waarschijnlijk gemaakt dat elke dierlijke cel is omgeven door een membraan met identieke structuur. In de Anglo-Amerikaanse literatuur spreekt men van 'unit-membranestructure'. Op doorsnede kan men in de elektronenmicroscoop aan zo'n membraan drie lagen onderscheiden: twee donkere lagen ter dikte van ongeveer 20 Å, ter weerszijden van een lichte die ongeveer 55 Å dik is. De totale membraandikte bedraagt dus 75 Å.Zowel de dendriet als het axon met zijn myelineschede vertonen eenzelfde membraanstructuur en ook de sandwichopbouw van het membraan is waarneembaar.
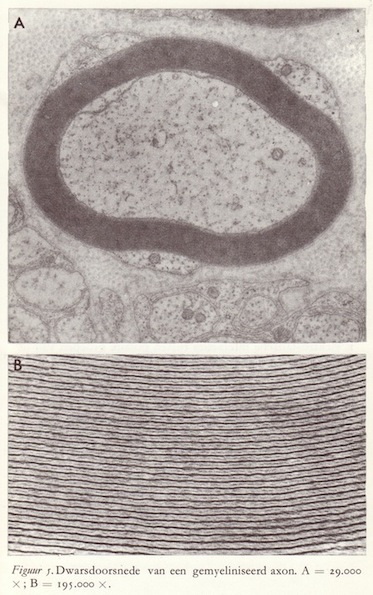
De opbouw van de myelineschede, die laag van vet-achtige stof die, slechts onderbroken door de insnoeringen van Ranvier, het axon isolerend omhult, kan het best begrepen worden aan de hand van wat zojuist over de membraanopbouw werd opgemerkt. De perifere zenuwvezels zijn omhuld door een tweede schede, het neurilemma, ook wel de 'schede van Schwann' genoemd. Het zijn nu de cellen waaruit deze schede bestaat, de zgn. Schwann-cellen, die verantwoordelijk zijn voor de aanwezigheid van de myelineschede. We kunnen zeggen dat, geschematiseerd, de myelineschede bestaat uit stijf opgerolde lagen van dubbele Schwann-celmembranen.
Het mechanisme van de myelinevorming in het centrale zenuwstelsel waar de Schwann-cellen afwezig zijn, wordt overgenomen door bepaalde gliacellen, nl. de oligodendroglia. Een gemyeliniseerd axon kan van enkele tot vele tientallen myelinelamellen bezitten. Elk van die lamellen bestaat uit twee bimoleculaire lipide-lagen, elk ongeveer 55 Å dik, gesandwiched tussen 20-30 Å-dikke eiwitlagen. De zich herhalende unit is ongeveer 150-160 Å dik. (18)
Moleculaire membraanstructuur
 Er zijn een aantal neurologische ziektebeelden bekend waarbij, zonder dat de chemische samenstelling van de myelinelamellen verandert, toch de regelmaat van de rangschikking van de lamellen verstoord wordt.
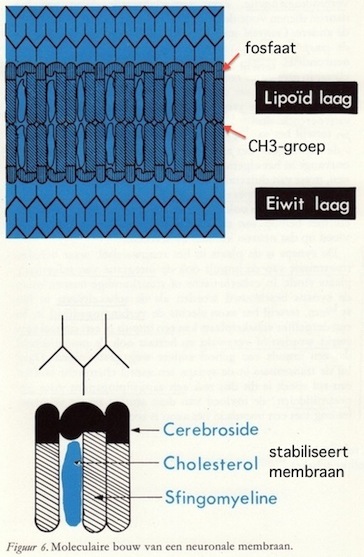
Er zijn een aantal neurologische ziektebeelden bekend waarbij, zonder dat de chemische samenstelling van de myelinelamellen verandert, toch de regelmaat van de rangschikking van de lamellen verstoord wordt.Ook wat betreft de chemische samenstelling van de myeline is de laatste jaren veel tot klaarheid gebracht. Myeline bestaat uit vetten, eiwitten, polysacchariden, zouten en water. De vetmoleculen zijn over het algemeen langgerekte structuren die ongeveer 26-28 Å meten en waarvan de kop uit een fosfaatmolecuul bestaat en de staart uit een CH3-groep. Elke membraaneenheid bestaat uit twee rijen vetmoleculen die a.h.w. staart aan staart liggen. Het centrale deel, dat dus bestaat uit niet-polaire CH2-groepen, wordt hydrofoob genoemd; het perifere deel is hydrofiel en bestaat uit polaire fosfaatgroepen.
De dikte van de lipidelagen is zo groot, dat we ons gemakkelijk kunnen voorstellen dat de specifieke myeline-moleculen zoals sfingomyeline, de cerebroside- en de cholesterolmoleculen in de lengte gerangschikt zijn. Zoals in detail in figuur 6 is weergegeven, staan deze moleculen in een bijzondere relatie tot elkaar.
De volgende factoren bepalen de stabiliteit van het menbraan:
a. elektrostatische interacties tussen polaire groepen van de vetten en de daaraan grenzende eiwitmoleculen;
b. de Van-der-Waals-krachten tussen de CH2-paren.
Wat zijn nu die mysterieuze Van-der-Waals-krachten en waarom is hun betekenis voor de moleculaire biologie zo groot? We kunnen ze definiëren als aantrekkingskrachten die ontstaan door het samenspel van elektrisch geladen systemen die polariseerbaar zijn. Volgens fysici is hun wijze van ontstaan het enige mechanisme dat zij zich kunnen voorstellen, waarbij specifieke intermoleculaire krachten eigenschappen vertonen die in de levende natuur blijken te bestaan. Dit uiterst complexe systeem is dus de basis waarop de impulsen langs de axonen getransporteerd worden. (20)
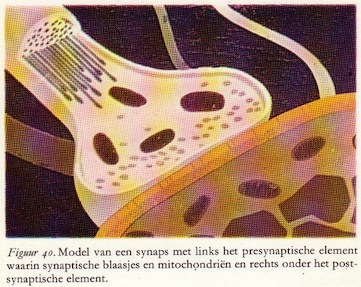
Schakelplaats in het zenuwstelsel: de synaps
 Het axon vervult in het perifere en centrale zenuwstelsel een verbindingsfunctie. Zeer gespecialiseerde delen van het neuron dienen voor de prikkeloverdracht van de ene cel naar de andere. Centraal staat hier de rol die wordt gespeeld door de synaps. Dit is een interneuronale verbinding waar twee neurondelen, elk behorend tot een verschillend neuron, met elkaar in zeer nauw contact treden. De neurondelen gaan niet in elkaar over, maar ze worden gescheiden door een zeer smalle ruimte: de synaptische kloof of spleet.
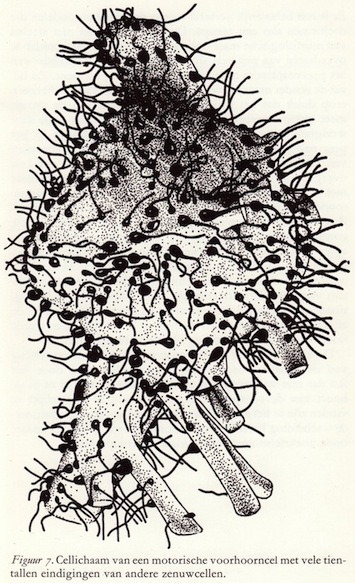
Het axon vervult in het perifere en centrale zenuwstelsel een verbindingsfunctie. Zeer gespecialiseerde delen van het neuron dienen voor de prikkeloverdracht van de ene cel naar de andere. Centraal staat hier de rol die wordt gespeeld door de synaps. Dit is een interneuronale verbinding waar twee neurondelen, elk behorend tot een verschillend neuron, met elkaar in zeer nauw contact treden. De neurondelen gaan niet in elkaar over, maar ze worden gescheiden door een zeer smalle ruimte: de synaptische kloof of spleet.Het impuls-aanvoerende deel van de synaps - de zender - wordt het presynapstische element genoemd en het impuls-ontvangend deel - de ontvanger - het postsynaptische element. Eén enkel neuron ontvangt in het algemeen prikkels van duizenden en tienduizenden andere neuronen. Het receptieve oppervlak van de zenuwcellen (cellichaam en dendrieten) is dan ook dicht bezet met axon-eindigingen, die elk met behulp van elektrische signalen hun specifieke invloed op dat neuron kunnen uitoefenen.
De synaps is de plaats in het zenuwstelsel, waar behalve transmissie van de impuls ook de integratie van informatie plaatsvindt. In cybernetische of stuurkundige termen moet de synaps worden beschouwd als de schakelplaats in het systeem, terwijl het axon de verbindingsdraad is die een impuls voortgeleidt. In een dergelijke schakelplaats kan een impuls of een signaal versterkt worden of verzwakt en bestaat ook de mogelijkheid dat een impuls een geheel andere weg gaat volgen.
Daar bij de transmissie in de synaps een aantal chemische stoffen een rol speelt, is dit ook een aangrijpingspunt voor geneesmiddelen; de invloed van deze stoffen op de voortgeleiding van een impuls in het axon is zeer gering. (22)
Unieke betekenis van neurotransmitters
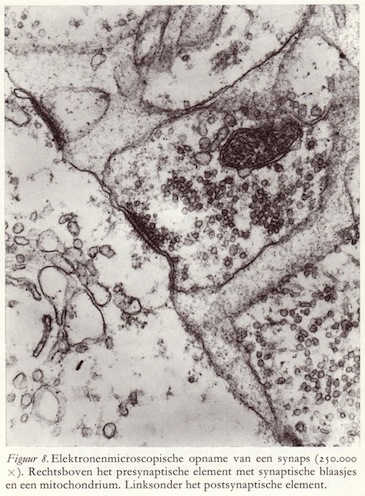 Er is een belangrijk verschil tussen de twee neurondelen die deelnemen aan een synaps. De verschillen zijn niet slechts van morfologische maar ook van fysiologische aard, omdat de overdracht van prikkels altijd in één richting geschiedt: van het presynaptische naar het postsynaptische element. Zo bevat de zender meer mitochondriën dan de ontvanger, hetgeen erop duidt dat het prikkel-uitsturende deel van de synaps meer energie verbruikt.
Er is een belangrijk verschil tussen de twee neurondelen die deelnemen aan een synaps. De verschillen zijn niet slechts van morfologische maar ook van fysiologische aard, omdat de overdracht van prikkels altijd in één richting geschiedt: van het presynaptische naar het postsynaptische element. Zo bevat de zender meer mitochondriën dan de ontvanger, hetgeen erop duidt dat het prikkel-uitsturende deel van de synaps meer energie verbruikt.Veel belangrijker echter is de aanwezigheid van een groot aantal uniforme blaasjes (granulae) in het presynaptische deel. Deze blaasjes hebben alle een diameter van 400 tot 600 Å en komen altijd daar voor waar prikkels overgedragen moeten worden, en alleen in het zendende, maar niet in het ontvangende element. Vandaar dat ze ook aangetoond zijn in zintuigcellen op plaatsen waar prikkeloverdracht plaatsvindt, op zenuwcellen en verder in de functionele contactplaatsen tussen zenuwcel en kliercel en zenuwcel en spiervezel. Hoewel de blaasjes er leeg uitzien, zijn ze dit in het geheel niet, want ze bevatten duizenden uiterst belangrijke moleculen: de zgn. transmitterstoffen (neurotransmitters), die verantwoordelijk zijn voor de prikkeloverdracht tussen neuronen onderling, tussen zintuigcellen en neuronen, en tussen neuronen en de effectoren zoals kliercellen en spieren. In het tweede en vierde hoofdstuk wordt hierop nog uitvoerig teruggekomen.
De in vergelijking tot het axon zo anders gerichte functie van de synaps wordt verder nog geïllustreerd door het feit dat zeer vele enzymen in hoge concentraties in of in de buurt van de synaps voorkomen, terwijl ze nauwelijks te vinden zijn in het axon. Een - misschien te ver gespecialiseerde - scheiding heeft in grote laboratoria wel geleid tot aparte onderzoekrichtingen: axonologie en synaptologie. (24)
terug naar de Inhoud
2. Elektrofysiologie
Onze kennis van de functie van het centrale en perifere zenuwstelsel is in de laatste jaren enorm toegenomen. Dit is voor een belangrijk deel te danken aan het voortschrijden van de techniek en in het bijzonder aan de ontwikkeling van elektronische apparatuur. De kleinste elektrische signalen, in de orde van grootte van miljoenste delen van een volt, die in zintuigcellen, zenuwcellen en spiervezels optreden, kunnen nu vastgelegd en geanalyseerd worden.
Alhoewel de activiteiten van de zenuwcellen een grote verscheidenheid van effecten teweegbrengen zoals willekeurige en onwillekeurige lichaamsbewegingen, bewuste en onbewuste denkprocessen, is de signaaloverdracht van relatief eenvoudige aard. Slechts twee typen potentialen, actiepotentialen (in het axon) en synaptische potentialen (in de synaps), zorgen voor de verwerking van miljoenen signalen, elk moment van de dag. Van het begin tot het einde van het leven is er een voortdurende elektrische activiteit in de hersenen van de mens.
De elektrofysiologie van het zenuwstelsel zal worden toegelicht met een eenvoudig voorbeeld: de reflexboog, waarin echter alle fysiologische fenomenen voorkomen die ook elders in het zenuwstelsel kunnen worden geregistreerd. De kleurenfoto's zijn ontleend aan een film, De Zenuwcel, die in samenwerking met de Stichting Film en Wetenschap te Utrecht geproduceerd werd naar tekeningen van C. van der Grond. (26)
Het zenuwstelsel: een informatieverwerkende rekenmachine
 Het zenuwstelsel is te beschouwen als een informatieverwerkende rekenmachine. Het ontvangt per tijdseenheid tienduizenden signalen, verwerkt deze, en geeft duizenden signalen af.
Het zenuwstelsel is te beschouwen als een informatieverwerkende rekenmachine. Het ontvangt per tijdseenheid tienduizenden signalen, verwerkt deze, en geeft duizenden signalen af.- De ontvangst van de signalen geschiedt door de receptorcellen van de zintuigorganen en
- de verwerking ervan door de complexe systemen van neuronale circuits in het perifere en centrale zenuwstelsel,
- terwijl de afgifte van de signalen voornamelijk paatsvindt aan de spieren en klieren.
De hoofdfuncties van het zenuwstelsel vallen uiteen in drie groepen:
1. Het integreert de verrichtingen van de elk op hun eigen functie gerichte organen op een doeltreffende manier; op een veel snellere manier ook dan ooit door stoffelijk vervoer via de bloedvaten of de lymfevaten mogelijk zou zijn. De organen worden op deze manier samengevat tot een eenheid van hogere orde: het individu.
2. Het individu kan dan op een doeltreffende en snelle manier reageren op veranderingen in de buitenwereld. Op deze wijze verdedigt het zijn integriteit en draagt bij tot het bestaan van de soort.
3. Bepaalde delen in het menselijke zenuwstelsel moeten worden beschouwd als de plaats waar, op overigens nog onbegrepen wijze, de verbinding wordt gelegd tussen geest en lichaam, en waar we het submicroscopische of moleculaire substraat moeten zoeken voor het abstracte denken.
Het opwekken en voortgeleiden van een impuls in het zenuwstelsel gaat gepaard met elektrische verschijnselen welke zich afspelen aan het membraan. In rust is het membraan elektrisch geladen, hetgeen polarisatie wordt genoemd (het membraan is gepolariseerd). De rustpotentiaal geeft dan aan: het potentiaalverschil tussen binnen en buitenkant van het membraan (de binnenkant negatief, de buitenkant positief). De geleiding in een zenuwcel heeft men wel vergeleken met een elektrische geleiding, maar de zenuwcel is - in tegenstelling tot een stroomdraad - ook zelf prikkelbaar en is dus niet alleen geleider, maar ook stroomgenerator (door een biochemische pomp die elektrisch geladen deeltjes, ionen, naar binnen of naar buiten pompt en zo de rustpotentiaal veroorzaakt).(28)
De reflexkring
 We kunnen ons nu afvragen hoe deze elektrische verschijnselen in de zenuwcel ontstaan en hoe ze langs de uitlopers worden geleid. Deze en andere vraagstukken (bijv. hoe de verandering in een elektrische potentiaal van de ene zenuwcel op de andere wordt overgebracht) zijn met behulp van elektronische micromethoden gedurende de laatste tiental jaren het onderwerp van intensieve studies geweest.
We kunnen ons nu afvragen hoe deze elektrische verschijnselen in de zenuwcel ontstaan en hoe ze langs de uitlopers worden geleid. Deze en andere vraagstukken (bijv. hoe de verandering in een elektrische potentiaal van de ene zenuwcel op de andere wordt overgebracht) zijn met behulp van elektronische micromethoden gedurende de laatste tiental jaren het onderwerp van intensieve studies geweest.De betreffende fenomenen zullen worden beschreven en geïllustreerd aan de hand van een eenvoudige schakeling van zenuwcellen, de reflexboog, ook wel reflexkring genoemd.
Deze bestaat in zijn meest algemene vorm uit:
1. Een sensorische (of afferente) zenuwvezel die zorgdraagt voor de inwerking van de impulsen uit de buitenwereld en voor het transport naar het centrale zenuwstelsel;
2. een of meer schakelcellen of interneuronen die prikkels, door de sensorische zenuwcel binnengebracht, verwerken en mededelen aan:
3. de motorische (of efferente) zenuwvezel. Deze brengt de informatie van het centrale zenuwstelsel over naar de spier voor de uitwerking.
Met betrekking tot de sensorische zenuwcel moet nog worden opgemerkt dat deze in vele gevallen niet in staat is om de veranderingen in onze omgeving of in ons lichaam zelf waar te nemen. In dergelijke gevallen is dan een zintuigcel aan het begin van de reflexboog opgenomen. Deze voert een transformatie uit, waardoor de veranderingen in de buiten- of binnenwereld worden omgezet in dusdanige elektrische verschijnselen, dat zij aan de sensorische zenuwcel kunnen worden meegedeeld. Deze reflexbogen zijn de pijlers waarop al het gebeuren in het zenuwstelsel berust.
De verschillende elektrische verschijnselen die in de onderscheiden delen van de reflexboog optreden, zullen nu bespreking vinden. (30)
Informatie-overdracht in een reflexkring
 Op zijn weg door de reflexboog doorloopt de prikkel de volgende stadia:
Op zijn weg door de reflexboog doorloopt de prikkel de volgende stadia:a. receptie van de prikkel in een zintuigcel;
b. prikkelgeleiding langs het membraan van de afferente zenuwvezel;
c. overdracht van de prikkel van de ene zenuwcel op de andere in de synaps;
d. prikkelgeleiding langs het membraan van de efferente zenuwvezel;
e. overdracht in de neuromusculaire synaps met als gevolg samentrekken van de desbetreffende spiervezel.
Een van de merkwaardigste feiten bij bestudering van de functie van het zenuwstelsel is wel dat de werking van dit orgaan, met al zijn ingewikkelde processen, waarschijnlijk is terug te voeren tot een aantal zeer elementaire elektrische verschijnselen zoals die besproken zullen worden aan de hand van het doorlopen van een prikkel in de reflexboog. De meest ingewikkelde mechanismen in het zenuwstelsel worden uitgevoerd door twee eenvoudige systemen van informatieverwerking en signaaloverdracht: de actiepotentiaal en de synaptische potentiaal. Als voorbeeld hebben we genomen een alledaags verschijnsel: het trappen op een kwal aan het strand.
De reflex: een onwillekeurige reactie
 Een op het strand wandelende badgast trapt op een kwal. Doordat zijn voetzool onverwachts met een koude, slijmerige massa in aanraking komt, treedt een complex van verschijnselen op. Zijn voet wordt werktuiglijk opgetild en pas later onderzoekt hij de oorzaak van de prikkel. Het eerste verschijnsel is een reflex die zich in het ruggemerg afspeelt, het tweede is een gewaarwording doordat de prikkel ook de schors van de grote hersenen bereikt.
Een op het strand wandelende badgast trapt op een kwal. Doordat zijn voetzool onverwachts met een koude, slijmerige massa in aanraking komt, treedt een complex van verschijnselen op. Zijn voet wordt werktuiglijk opgetild en pas later onderzoekt hij de oorzaak van de prikkel. Het eerste verschijnsel is een reflex die zich in het ruggemerg afspeelt, het tweede is een gewaarwording doordat de prikkel ook de schors van de grote hersenen bereikt.1. De eerste reactie is zinvol, ook al geschiedt zij zonder invloed van de wil. Langs afferente zenuwvezels bereiken de prikkels het sacrale deel van het ruggemerg. Van hier schakelen zij over door een of meerdere schakelcellen op efferente zenuwcellen. Langs de efferente zenuwvezels arriveren de prikkels in de spieren, waar zij een contractie teweegbrengen. Het zichtbare gevolg is een reflex: het automatische optillen van de voet. Hierbij worden niet alleen bepaalde spiergroepen geactiveerd, maar andere moeten door dezelfde prikkel geremd worden.
2. Ondertussen worden de prikkels ook door het ruggemerg naar de hersenen gezonden om daar bewust te worden gemaakt. Hierna volgt een bewuste reactie: het onderzoek naar de oorzaak van de prikkel en het nemen van verdere maatregelen.
Deze gescheiden processen volgen zo snel op elkaar dat het lijkt alsof we met één proces te doen hebben. Dit is mogelijk doordat de prikkels in het zenuwstelsel zich met grote snelheid langs de zenuwvezels voortplanten. Nadat de strandwandelaar heeft gezien dat hij op een kwal heeft getrapt, worden prikkels vanuit de motorische schors van de grote hersenen naar het ruggemerg gestuurd: een willekeurige reactie volgt dan op de onwillekeurige reactie. In het algemeen geldt dat een reflex erop is gericht de oorzaak van zijn ontstaan tegen te werken, zoals ook bij ons voorbeeld het geval is.
De weg die de prikkel van voetzool naar ruggemerg en van ruggemerg naar de beenspieren heeft gevolgd, zullen wij nu ook doorlopen. (34)
De receptoren-zintuigcellen
 Bij de zoogdieren en de mens vinden wij zintuigcellen - receptoren - die reageren op verschillende soorten prikkels.
Bij de zoogdieren en de mens vinden wij zintuigcellen - receptoren - die reageren op verschillende soorten prikkels.De zintuigcellen in het oog reageren op licht, die in het oor op geluid of op veranderingen in de evenwichtstoestand, die in de huid op druk, warmte, koude, pijn, etc. Alle zintuigcellen zijn onder natuurlijke omstandigheden speciaal gevoelig voor één bepaalde energiesoort en slechts gering of helemaal niet voor andere. De zintuigcellen registreren de specifieke verandering in bijv. fysisch of chemisch opzicht, die zij van de buitenwereld opvangen, maar deze energie wordt niet rechtstreeks op het uiteinde van de sensibele zenuwcel overgebracht. De zintuigcel transformeert als het ware de energie waarvoor deze cel gevoelig is: licht-energie bijvoorbeeld, opgevangen door de zintuigcellen in het oog, wordt door sensibele zenuwcellen als elektrische veranderingen naar het centrale zenuwstelsel vervoerd.
De veranderingen in de buitenwereld, zoals deze aan de huid worden medegedeeld, brengen veranderingen teweeg in de zintuigcellen in de huid of in het onderhuidse bindweefsel en deze op hun beurt geven de informatie door aan de uiteinden van de sensibele zenuwen. De beide figuren hiernaast geven schematisch en sterk vergroot de huid onder de bal van de voet weer: de donkerbruine onderrand stelt de bloedarme opperhuid voor, daarboven de bloedrijke lederhuid; het eivormige lichaampje links is een zintuigcel die gevoelig is voor drukverschillen: een mechanoreceptor, het lichaampje van Vater Pacini geheten. Het bestaat uit een aantal concentrische lamellen, ook wel schillen genoemd. De witte draad die in de lengte-as van de receptor verloopt, is het uiteinde van de sensibele zenuw. Er naast is een rijkvertakte eindiging van een andere sensibele zenuw getekend.
De figuur 16 (links) geeft aan de huid in rust, en 16 (rechts) de huid wanneer daarop een bepaalde druk wordt uitgeoefend zoals bij de aanraking van de voet met de kwal. (36)
Potentiaalverschil in de zintuigcel
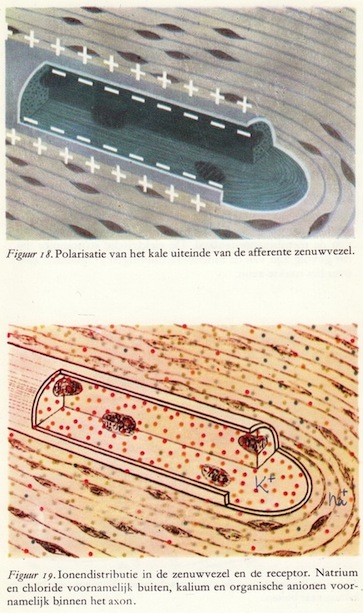 De figuren 18 en 19 zijn een sterkere vergroting van het binnenste deel van het tastlichaampje. Het uiteinde van de sensibele zenuw is 'naakt'. In de figuur is het 'naakte' zenuwuiteinde opengesneden, zodat we de binnenkant kunnen zien, wat natuurlijk in werkelijkheid niet het geval is. Het isolerende materiaal of de myelineschede dringt gedeeltelijk door in de receptor. Eén insnoering van Ranvier bevindt zich nog juist binnen de receptor. Het uiteinde van de zenuwvezel zelf is niet geïsoleerd.
De figuren 18 en 19 zijn een sterkere vergroting van het binnenste deel van het tastlichaampje. Het uiteinde van de sensibele zenuw is 'naakt'. In de figuur is het 'naakte' zenuwuiteinde opengesneden, zodat we de binnenkant kunnen zien, wat natuurlijk in werkelijkheid niet het geval is. Het isolerende materiaal of de myelineschede dringt gedeeltelijk door in de receptor. Eén insnoering van Ranvier bevindt zich nog juist binnen de receptor. Het uiteinde van de zenuwvezel zelf is niet geïsoleerd.Ook dit uiteinde vertoont een potentiaalverschil tussen binnen- en buitenkant van het membraan. Binnen het axon worden hoofdzakelijk kaliumionen (K+) gevonden (aangegeven met rode punten) en organische ionen (aangegeven met gele punten). Buiten het membraan van het axon, maar nog binnen de receptor, vinden we voornamelijk natrium- (Na+) en chloorionen (Cl-) (aangegeven door resp. blauwe en groene stippen). Concentratieverschillen en een specitieke doorlaatbaarheid van het membraan voor ieder der diverse ionen veroorzaken een potentiaalverschil tussen de binnen- en buitenkant van het memhraan in rusttoestand: de buitenkant positief, de binnenkant negatief. Deze rustpotentiaal heeft een waarde van ongeveer -60 mV.
Ook zijn nog een viertal ovaalvormige orgaantjes getekend. Dit zijn mitochondriën die een halve tot enkele μ's lang zijn. In een grote zenuwcel bevinden zich 5000 tot 10.000 van deze mitochondriën. Elk van deze structuurtjes, zoals reeds aangestipt werd op blz. 16, kan vergeleken worden met een krachtcentrale waarvan de belangrijkste taak is de energie te leveren voor de functie van de cel.
De elektrische lading van het membraan van het uiteinde van een sensibele vezel in de receptor wordt in rust gehandhaafd op ongeveer -60 mV. Wanneer echter druk op de huid wordt uitgeoefend, waardoor de lamellen in de receptor langs elkaar gaan schuiven, treden er veranderingen op in de rustpotentiaal. De mechanische prikkel wordt omgezet in een elektrisch signaal. (38)
Ionenverschuivingen in de zintuigcel
De verschuivingen van de lamellen van de drukreceptor (mechanische verandering) veroorzaken verplaatsingen van ionen door het 'naakte' membraan van het axon. Eerst stroomt een geringe hoeveelheid natriumionen van buiten naar binnen. De blauwe pijl in de figuren 20 en 21 geeft deze stroom van natriumionen aan.
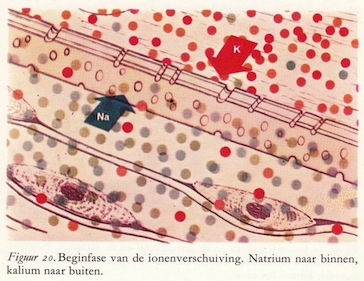
Direct daarna volgt een verplaatsing van een geringe hoeveelheid kaliumionen van binnen het axon door het membraan naar buiten. Deze uitstroom van kaliumionen is aangegeven door de rode pijl. De verschuivingen van de lamellen, door de druk op de receptor teweeggebracht, hebben de doorlaatbaarheid van het membraan voor de ionen veranderd. De ionenverplaatsingen veroorzaken een kleine depolarisatie of ontlading van het membraan van het uiteinde van het axon.
Dit kunnen we op de oscilloscoop aflezen als een verandering van de membraanpotentiaal. De elektrische stroompjes die het gevolg zijn van de ionenverschuivingen verplaatsen zich over het naakte axon. (40)
Dit kunnen we op de oscilloscoop aflezen als een verandering van de membraanpotentiaal. De elektrische stroompjes die het gevolg zijn van de ionenverschuivingen verplaatsen zich over het naakte axon. (40)
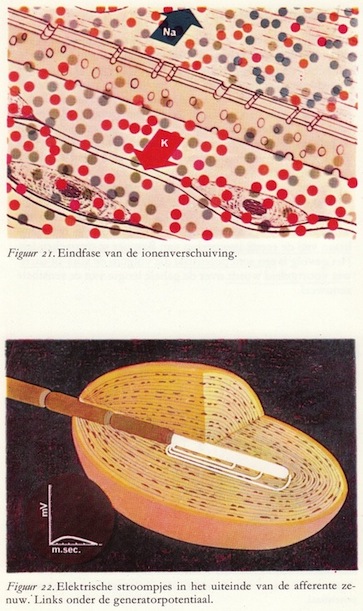
Generatorpotentialen
De kleine potentiaalveranderingen in het membraan van het axon worden generatorpotentialen genoemd omdat zij uiteindelijk een totale ontlading van het membraan veroorzaken, waardoor een actiepotentiaal voortgeleid kan worden.

Als de prikkel te zwak is (fig. 25) zijn de generatorpotentialen slechts enkele mV groot en te gering om de drempelwaarde te overschrijden. Wordt de druk op de receptor groter en daardoor de prikkel sterker, dat nemen de ionenverschuivingen toe, de generatorpotentialen worden groter en ook de elektrische stroompies nemen in intensiteit toe (fig. 24).
Bij een bepaalde prikkelsterkte wordt de drempelwaarde bereikt: de generatorpotentialen hebben dan een maximale waarde. De elektrische stroompjes zijn nu zo krachtig geworden dat zij in staat zijn de drempelwaarde van het membraan van de eerste insnoering van Ranvier te overschrijden.
Het gevolg is een actiepotentiaal, die volgens de alles-of-nietswet wordt voortgeleid over de gehele lengte van de sensibele zenuwcel. (42)
Bij een bepaalde prikkelsterkte wordt de drempelwaarde bereikt: de generatorpotentialen hebben dan een maximale waarde. De elektrische stroompjes zijn nu zo krachtig geworden dat zij in staat zijn de drempelwaarde van het membraan van de eerste insnoering van Ranvier te overschrijden.
Het gevolg is een actiepotentiaal, die volgens de alles-of-nietswet wordt voortgeleid over de gehele lengte van de sensibele zenuwcel. (42)
De actiepotentiaal 1
 Als voorbeeld nemen we een niet-gemyeliniseerde, enkele zenuwvezel, die zich bevindt in een niet-geleidend medium. Op de volgende bladzijde zullen we dan dieper ingaan op de speciale verschijnselen die optreden in de gemyeliniseerde zenuwvezels.
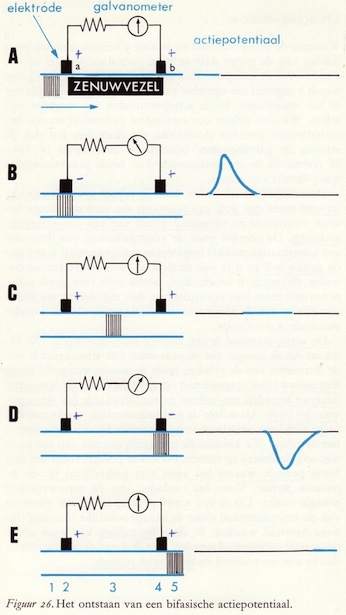
Als voorbeeld nemen we een niet-gemyeliniseerde, enkele zenuwvezel, die zich bevindt in een niet-geleidend medium. Op de volgende bladzijde zullen we dan dieper ingaan op de speciale verschijnselen die optreden in de gemyeliniseerde zenuwvezels.Een paar registratie-elektroden wordt aan het axon bevestigd en met behulp van een galvanometeropstelling (ampèremeter, voor het meten van elektrische stromen) wordt de elektrische reactie van het axon op een kunstmatige prikkel waargenomen. We registreren dus de uitslagen van de galvanometer. Gesteld dat we een zenuwvezel bij het punt 1 (links onderaan) geprikkeld hebben. Wanneer de actiepotentiaal die in de zenuw wordt opgewekt, positie 2 op het axon bereikt, zien we een uitslag van de galvanometer. De elektrische lading van het membraan van de zenuwvezel verandert blijkbaar. We spreken van ontladen of depolariseren. De meter slaat dan bijv. naar links uit, hetgeen door een opwaartse lijn in het rechtergedeelte van de tekening (onder 'actiepotentiaal') wordt aangegeven.
Door deze plaatselijke ontlading van het membraan kan elektrode a beschouwd worden in contact te zijn met het negatief geladen binnenste van het axon. We hebben nl. in de inleiding op blz. 28 gezien dat in de rusttoestand het axon, en dus ook de hele zenuwcel, aan de binnenkant negatief en aan de buitenkant positief geladen is. De elektrode b is in contact met het nog positief geladen buitenste van het gedepolariseerde membraan. Vandaar het tijdelijk optreden van een potentiaalverschil tussen de elektroden en dus ook het verschijnsel dat er gedurende korte tijd stroom loopt in de galvanometer en wel, zoals in ons geval in de tekening is weergegeven, van rechts naar links. De lijn in het rechtergedeelte van de figuur is opwaarts getekend daar het onder neurofysiologen gebruikelijk is om hun waarnemingsinstrumenten zo te schakelen dat een opwaartse lijn in een figuur aangeeft dat de elektrode, die verbonden is met het actieve deel van de zenuw, meer negatief wordt ten opzichte van de inactieve elektrode. (44)
De actiepotentiaal 2
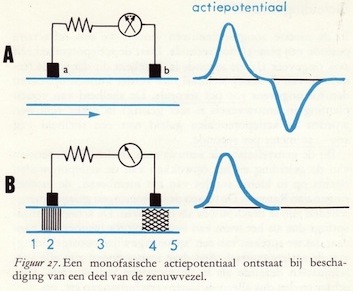 Wanneer de actiepotentiaal nu positie 3 bereikt, zien we geen uitslag van de meter daar er geen potentiaalverschil tussen a en b is. Als de actiepotentiaal echter aankomt in positie 4 dan wordt b negatief ten opzichte van a: de stroom draait dus om in het instrument. Beide actiepotentialen zijn negatief van teken. We zien echter een opwaartse potentiaal en een benedenwaarts gerichte potentiaal, dit door het feit dat de stroom de galvanometer beurtelings binnengaat of links of rechts. In de rechtertekening zijn beide potentialen het spiegelbeeld van elkaar.
Wanneer de actiepotentiaal nu positie 3 bereikt, zien we geen uitslag van de meter daar er geen potentiaalverschil tussen a en b is. Als de actiepotentiaal echter aankomt in positie 4 dan wordt b negatief ten opzichte van a: de stroom draait dus om in het instrument. Beide actiepotentialen zijn negatief van teken. We zien echter een opwaartse potentiaal en een benedenwaarts gerichte potentiaal, dit door het feit dat de stroom de galvanometer beurtelings binnengaat of links of rechts. In de rechtertekening zijn beide potentialen het spiegelbeeld van elkaar.De actiepotentiaal in het axon is echter geen elektrische stroom maar een golf van activiteit die zich snel langs het axon voortplant en vergezeld wordt van een potentiaalverandering. De energie voor de voortgeleiding van deze met een kunstmatige prikkel opgewekte actiepotentiaal komt van de zenuw zelf en niet van de prikkel. Wordt het axonstukje onder elektrode b beschadigd, zodat zich hier geen actiepotentiaal meer kan voortplanten, dan registreren we alleen een uitslag van de galvanometer als de actiepotentiaal onder elektrode a doorloopt.
De actiepotentiaal is een alles-of-niets-fenomeen. Dit betekent dat de hoogte van de potentiaal niet afhankelijk is van de intensiteit van de prikkel, noch de manier van prikkeling. Een zenuwvezel reageert niet op een prikkel als de intensiteit daarvan beneden een zekere minimumwaarde (de drempelwaarde) blijft. Als echter de prikkelintensiteit daarboven ligt, volgt altijd een reactie en voor eenzelfde axon is deze reactie steeds gelijk.
We kunnen de prikkelfrequentie van het axon niet ongelimiteerd opvoeren. Na iedere prikkel volgt een zeer korte periode waarin het axon niet prikkelbaar is: de refractaire periode. Voor het ontladen van de zenuwvezel is energie nodig. Dit is een actief proces en na het passeren van de actiepotentiaal moet de oorspronkelijke toestand dus weer hersteld worden. In deze herstelfase kan geen actiepotentiaal voortgeleid worden. Het deel van de zenuwvezel is dan in wat we noemen de refractaire periode. (46)
Geleidingssnelheid
 In de meeste zoogdierzenuwen vinden we een refractaire periode van plm. 1/1000 seconde. Daar de actiepotentiaal zelf ook ongeveer 1/1000 seconde is, betekent dit dat we de frequentie van de actiepotentiaal niet verder kunnen opvoeren dan tot ongeveer 500 per seconde. De snelheid van voortplanting in zenuwvezels is niet gering: in dikke zenuwen worden de actiepotentialen geleid met een snelheid van 100-120 meter per seconde.
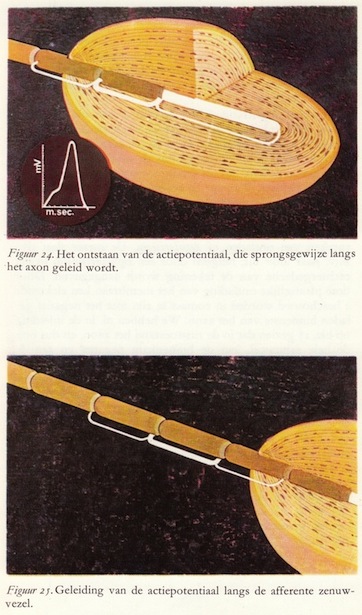
In de meeste zoogdierzenuwen vinden we een refractaire periode van plm. 1/1000 seconde. Daar de actiepotentiaal zelf ook ongeveer 1/1000 seconde is, betekent dit dat we de frequentie van de actiepotentiaal niet verder kunnen opvoeren dan tot ongeveer 500 per seconde. De snelheid van voortplanting in zenuwvezels is niet gering: in dikke zenuwen worden de actiepotentialen geleid met een snelheid van 100-120 meter per seconde.Bij de gemyeliniseerde zenuwvezels treden alle processen van de geleiding en het opwekken van de actiepotentialen slechts op in kleine stukjes van het membraan: tussen de insnoeringen van Ranvier. De tussen de insnoeringen gelegen mergscheden zijn te beschouwen als isolatoren. De actiepotentiaal springt dus als het ware van insnoering tot insnoering, vandaar dat we spreken van een 'sprongsgewijze voortgeleiding' van de actiepotentiaal. Aan deze insnoeringen zijn alle verschijnselen dezelfde als in de mergloze zenuwvezels. Hier treden dus alleen de ionenverschuivingen etc. op. (48)
De rustpotentiaal van de zenuwcel
 We gaan nu weer terug naar de rustpotentiaal van het axon en vragen ons af hoe deze potentiaal bepaald wordt. Het blijkt dat ook hier belangrijke verschillen bestaan in de concentraties van een aantal ionen aan beide kanten van het membraan. De concentratie van chloor- en natriumionen is buiten groter dan binnen, maar de concentratie van kaliumionen is binnen veel groter dan buiten. Deze situatie zijn we ook al tegengekomen bij het naakte uiteinde van het axon in de receptor, het lichaampje van Vater Pacini.
We gaan nu weer terug naar de rustpotentiaal van het axon en vragen ons af hoe deze potentiaal bepaald wordt. Het blijkt dat ook hier belangrijke verschillen bestaan in de concentraties van een aantal ionen aan beide kanten van het membraan. De concentratie van chloor- en natriumionen is buiten groter dan binnen, maar de concentratie van kaliumionen is binnen veel groter dan buiten. Deze situatie zijn we ook al tegengekomen bij het naakte uiteinde van het axon in de receptor, het lichaampje van Vater Pacini.Tijdens de actiepotentiaal wordt de binnenkant van het membraan niet alleen minder negatief, maar zelfs positief en meet dan gedurende korte tijd ongeveer + 20 mV, terwijl de potentiaal van het membraan in rust ongeveer -60 mV is.
Deze omkering van de potentiaal gaat gepaard met ionenverschuivingen. We kunnen aan de actiepotentiaal twee delen onderscheiden: een opstijgend deel en een afdalend deel.
1. Gedurende het opstijgende deel van de actiepotentiaal, waarin de lading aan de binnenkant van het membraan omdraait van negatief naar positief, wordt het membraan zeer specitiek doorgankelijk voor natriumionen. We zien dan een instroom van die ionen door het membraan van buiten naar binnen, zoals in de figuur is aangegeven.
Tijdens het afdalende deel van de actiepotentiaal neemt de doorlaatbaarheid van het membraan voor kaliumionen sterk toe, met gevolg een uitstroom van kaliumionen van binnen door het membraan naar buiten (de rode pijl in de figuur).
Tengevolge van de veranderde potentiaal in de insnoering van Ranvier, waar de actiepotentiaal opgewekt wordt, treden in dat deel elektrische stroompjes op die lopen van insnoering tot insnoering. Deze insnoeringen van Ranvier bevinden zich in een zenuwvezel van 10μ diameter op een afstand van ongeveer 700μ van elkaar. Men zou dus deze sprongsgewijze voortgeleiding van de actiepotentiaal kunnen opvatten als een economisch principe van deze zenuwvezels. (50)
Codering in een axon
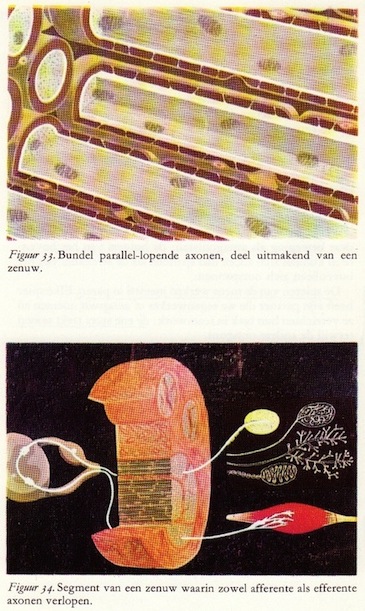 Aparte axonen, zoals in de vorige figuren weergegeven, komen in het zenuwstelsel van zoogdieren zo in het lichaam niet voor. Ze lopen altijd in bundels en bundels van axonen worden een zenuw genoemd.
Aparte axonen, zoals in de vorige figuren weergegeven, komen in het zenuwstelsel van zoogdieren zo in het lichaam niet voor. Ze lopen altijd in bundels en bundels van axonen worden een zenuw genoemd.Hoe is nu zo'n zenuw opgebouwd? In het algemeen slechts uit twee elementen: in de eerste plaats de axonen met hun myelinescheden en in de tweede plaats bindweefsel, dat mede zorgt voor een efficiënte isolatie van de axonen. Door dat bindweefsel worden de parallel aan elkaar lopende axonen op hun plaats gehouden en voor beschadiging behoed. Het vormt een samenhangend geheel en bestaat uit bindweefselcellen, bindweefselvezels en bindweefsellamellen. De twee laatste zijn het die de zenuw zijn stevigheid geven.
Naar zijn lokalisatie en structuur verdelen wij het bindweefsel in endoneurium, perineurium en epineurium. Het zeer fijn-vezelige endoneurium omhult direct het neurilemma van het axon. Het perineurium, bestaande uit dunne bindweefsellamellen, ligt tussen de axonen in en groepeert ze in bundels. Het epineurium tenslotte is de stevige bindweefselschede die de zenuw als geheel omhult en hem zijn peesachtige taaiheid verleent.
Een enkele zenuwvezel kan niets anders doen dan aparte actiepotentialen geleiden. Voor een zenuwvezel bestaat er dus maar één mogelijkheid tot transport van informatie: de codering geschiedt door middel van impulsfrequenties.
Voor een zenuw, dus voor een groot aantal parallel lopende zenuwvezels, bestaat er een extra coderingsmogelijkheid, nl. het aantal zenuwvezels dat bij de informatie-overdracht betrokken is; we zouden kunnen spreken van amplitudomodulatie.
Bij de gewaarwording van zelfs maar de geringste zintuiglijke prikkel zijn vele receptoren en een groot aantal zenuwvezels betrokken - het signaal in een enkele zenuwvezel heeft geen betekenis, maar het zeer complexe signaal in de bundel zenuwvezels kan tot een zinvolle gewaarwording verwerkt worden. (52)
Antagonisten en synergisten
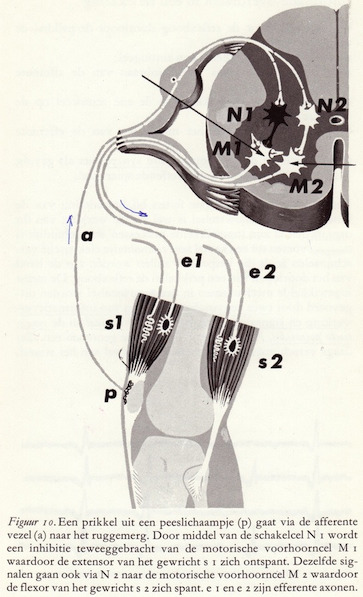
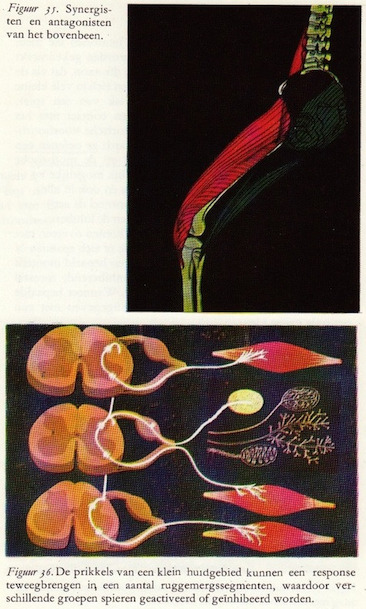 In de beginsituatie, waarbij de strandwandelaar op de kwal trapte en reflectoir zijn voet optilde, gingen sensibele prikkels langs de afferente zenuwvezels naar het ruggemerg. In de grijze stof springen de prikkels over een of meer schakelcellen over op de motorische zenuweel. Deze is het uitvoerende deel van de reflex. Zelfs bij een eenvoudige reflex zijn een groot aantal spieren betrokken, waarbij we als voorbeeld nemen de spiergroepen die aan de voorkant en achterkant van het dijbeen gelegen zijn en die zorgen voor de buiging en strekking van het bovenbeen. In figuur 55 zijn schematisch als rood aangemerkt de spieren die het bovenbeen buigen (voorbeelden: m. iliopsoas en m. rectus femoris) en als groen de spieren die het bovenbeen strekken (voorbeelden: m. gluteus maximus en m. biceps femoris). De beide groepen werken bij deze beweging als antagonisten: de buigers van het bovenbeen kunnen zich alleen dan spannen als de strekkers van het bovenbeen zich ontspannen.
In de beginsituatie, waarbij de strandwandelaar op de kwal trapte en reflectoir zijn voet optilde, gingen sensibele prikkels langs de afferente zenuwvezels naar het ruggemerg. In de grijze stof springen de prikkels over een of meer schakelcellen over op de motorische zenuweel. Deze is het uitvoerende deel van de reflex. Zelfs bij een eenvoudige reflex zijn een groot aantal spieren betrokken, waarbij we als voorbeeld nemen de spiergroepen die aan de voorkant en achterkant van het dijbeen gelegen zijn en die zorgen voor de buiging en strekking van het bovenbeen. In figuur 55 zijn schematisch als rood aangemerkt de spieren die het bovenbeen buigen (voorbeelden: m. iliopsoas en m. rectus femoris) en als groen de spieren die het bovenbeen strekken (voorbeelden: m. gluteus maximus en m. biceps femoris). De beide groepen werken bij deze beweging als antagonisten: de buigers van het bovenbeen kunnen zich alleen dan spannen als de strekkers van het bovenbeen zich ontspannen.De spieren werken meestal in paren. Elke spier heeft zijn partner die we tegenwerker of antagonist noemen en ze verrichten hun taak in teamwork: de ene spier trekt samen terwijl de andere ontspant.
De inkomende sensibele prikkels hebben in dit geval een tweeledige taak: het activeren of exciteren van de motorische zenuwen die de buigers innerveren en tevens het remmen of inhiberen van de motorische zenuwen die de strekkers van het bovenbeen innerveren. Figuur 56 geeft dit weer. De prikkels vanuit de receptor komen in het middelste segment binnen.
Van de drie spieren die getekend zijn, wordt bijv. de middelste (geïnnerveerd door motorische zenuwvezels uit het middelste segment) geactiveerd, terwijl de bovenste en onderste (geïnnerveerd door motorische voorhoorncellen in het bovenste en onderste ruggemergsegment) geremd worden.
Excitatie en inhibitie zijn de twee fysiologische grondbeginselen. Deze processen spelen zich af in het membraan van de motorische zenuwcel. We zullen nu iets dieper ingaan op de fysiologische eigenschappen van dit neuron. (54)
De motorische zenuwcel: activerende of exciterende synapsen
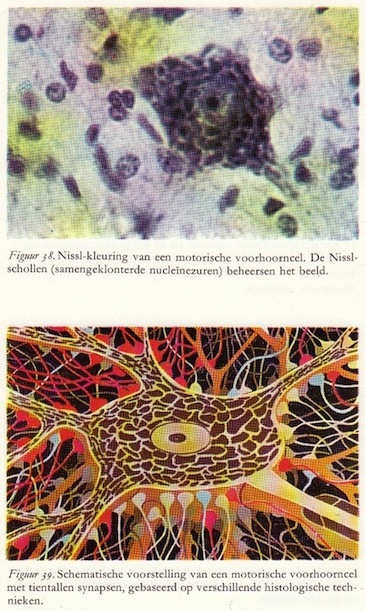 Motorische voorhoorncellen zijn grote neuronen die meer dan 100μ in diameter kunnen zijn. Ze worden gekenmerkt door een uitgebreid dendrietenstelsel en één dik axon, dat door de voorwortel loopt in de perifere zenuwen en zich in vele kleine vertakkinkjes uitbreidt op het oppervlak van een spier.
Motorische voorhoorncellen zijn grote neuronen die meer dan 100μ in diameter kunnen zijn. Ze worden gekenmerkt door een uitgebreid dendrietenstelsel en één dik axon, dat door de voorwortel loopt in de perifere zenuwen en zich in vele kleine vertakkinkjes uitbreidt op het oppervlak van een spier.Duizenden uiteinden van axonen maken contact met het cellichaam en de dendrieten van de motorische voorhoorncellen. De synapsen zijn van tweeërlei aard: ze oefenen een exciterende of een inhiberende invloed op de motorische voorhoorncel uit. Teneinde een zo duidelijk mogelijke voorstelling hiervan te geven, zijn in figuur 59 en ook in alle volgende figuren van de motorische voorhoorncel de activerende of exciterende synapsen rood getekend en de inhiberende of remmende synapsen blauw. Deze kleuren komen overeen met de kleuraanduiding in de spiergroepen die of zich spannen of zich ontspannen, zoals in figuur 35. Op een bepaald moment zijn niet alle synapsen exciterend of inhiberend, meestal functioneert slechts een klein percentage. Wanneer bepaalde synapsen niet actief zijn, worden ze aangegeven met een neutrale grijze kleur. (56)
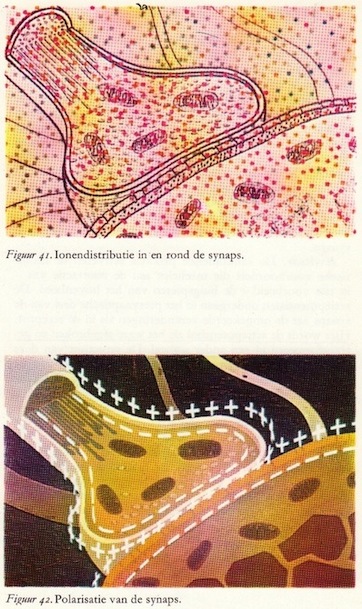
Ionen en rustpotentiaal van de synaps
Lichten we de synaps in een veel sterkere vergroting uit de tekening, dan blijkt dat ook hier dezelfde verschillen in ionenconcentraties aanwezig zijn als bij het axondeel in de receptor en de insnoeringen van Ranvier.
Zowel in het pre- als in het postsynaptische membraan vinden we een hoge concentratie aan kalium- en organische anionen en een lage concentratie aan natrium- en chloorionen, terwijl in de ruimte buiten deze twee neurondelen, de extracellulaire ruimte, een hoge concentratie aan natrium- en chloorionen en een lage concentratie aan kaliumionen wordt gevonden. De gekleurde balletjes hebben dezelfde betekenis als bij de figuren van de receptor en de knoop van Ranvier. De verschillen in ionenconcentraties en ionendoorlaatbaarheid van de membranen geven wederom aanleiding tot potentiaalverschillen, zowel in het pre- als in het postsynaptische element. In beide gevallen is de binnenkant negatief t.o.v. de buitenkant en ook hier een rustpotentiaal die ongeveer 60-70 mV bedraagt. (58)
Prikkel van receptor naar synaps: excitatie en inhibitie
 Nu we de basis hebben leren kennen waarop de prikkeloverdracht van de ene zenuwcel op de andere zenuwcel plaatsvindt, gaan we weer terug naar het afferente of aanvoerende deel van de reflexkring.
Nu we de basis hebben leren kennen waarop de prikkeloverdracht van de ene zenuwcel op de andere zenuwcel plaatsvindt, gaan we weer terug naar het afferente of aanvoerende deel van de reflexkring.De receptoren in de huid zorgden ervoor dat langs vele afferente vezels series prikkels de grijze stof in het ruggemerg bereiken. Deze komen over schakelcellen aan op het oppervlak van de motorische voorhoorncellen. Deze aankomende signalen kunnen nu twee verschillende processen teweegbrengen in de motorische voorhoorncellen: excitatie en inhibitie. De aankomende signalen zijn gelijk van aard: actiepotentialen van gelijke duur en gelijke hoogte, dus hierin moeten we het verschil niet zoeken; wel echter in de wijze waarop de aankomende prikkels overgedragen worden in de synaps.
Excitatie
De aankomende prikkels activeren de motorische voorhoorncel die meehelpt aan de contractie van - in ons voorbeeld - de buigspieren van het bovenbeen. De actiepotentialen ondergaan in het presynaptische deel van de synaps net de omgekeerde veranderingen als in de receptor.
Hier wordt de actiepotentiaal als het ware afgebroken en getransformeerd in een ander signaal. De structuur van deze twee zenuwuiteinden vertoont grote overeenkomst: in de receptor vonden we het 'naakte' deel van het axon en het presynaptische deel van het axon heeft eveneens een uiteinde dat zijn isolerende myelineschede verloren heeft.
Vervolgen we de actiepotentiaal op zijn weg langs de afferente zenuwvezel naar de synaps dan springt hij over van de laatste insnoering van Ranvier op het kale uiteinde van de vezel. Hier kan hij niet verder want, hoewel de afstand tussen het presynaptische en postsynaptische element in de synaps maar zeer klein is, kan deze elektrische prikkel toch niet van het ene neuron op het andere neuron overspringen. Het 'doodlopen' van die actiepotentiaal geeft een belangrijke verstoring in de elektrische eigenschappen van het presynaptische deel van de synaps. (60)
a. Excitatie door een transmitterstof
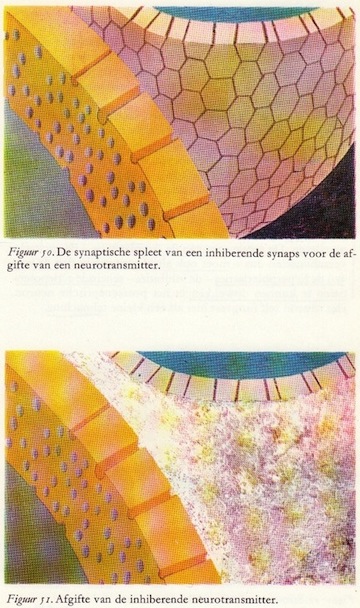
 Het aankomen van de actiepotentiaal in het presynaptische element, hier aangegeven met de rode kleur van excitatie, mobiliseert de aanwezige synaptische blaasjes (door depolarisatie). Deze worden geactiveerd en bewegen zich in groten getale naar het membraan, waar ze zich openen en de transmitterstof vrijmaken in de ruimte van de synaptische spleet.
Het aankomen van de actiepotentiaal in het presynaptische element, hier aangegeven met de rode kleur van excitatie, mobiliseert de aanwezige synaptische blaasjes (door depolarisatie). Deze worden geactiveerd en bewegen zich in groten getale naar het membraan, waar ze zich openen en de transmitterstof vrijmaken in de ruimte van de synaptische spleet.Het bovenste deel van figuur 45 geeft nog net een randje aan van het presynaptische element. Het gele membraan rechtsonder in de figuur is het postsynaptische element. De transmitterstof is in de synaptische spleet getekend en ziet eruit als wit vlokkig materiaal. Deze exciterende overdrachtstof verandert de doorlaatbaarheid van het postsynaptische membraan. Door deze verandering in de permeabiliteit treden ionenverschuivingen op, die wat hun karakter betreft vergelijkbaar zijn met de ionenverschuivingen in het axon tijdens de actiepotentiaal, maar veel geringer van omvang. Ook hier dus, zoals in figuur 46 is aangegeven, een instroom van natrium- en een uitstroom van kaliumionen.
Op welke wijze beïnvloeden nu deze ionenverschuivingen de activiteitstoestand van de motorische voorhoorncel? De elektrische verschijnselen die gepaard gaan met excitatie (hetzelfde geldt voor inhibitie) kunnen worden onderzocht met behulp van zeer dunne elektroden die in het binnenste van de motorische voorhoorncel gebracht worden. Daar de potentialen worden geregistreerd van het postsynaptische element, spreken we op de volgende bladzijden van postsynaptische potentialen. (62)
b. Excitatie door potentialen: depolarisatie
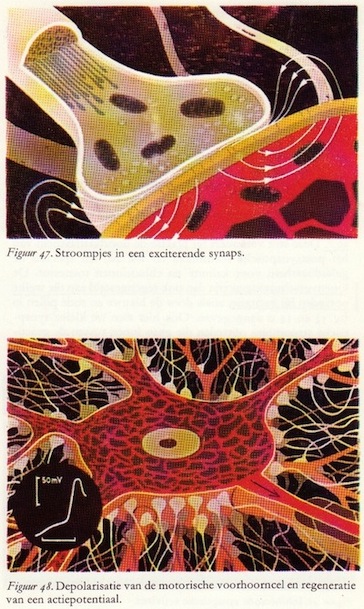 Doordat de doorlaatbaarheid van het membraan alleen zeer lokaal verandert, treden er kleine stroompjes op die door de synaps lopen (de richting hiervan is aangegeven in figuur 47), en zich steeds verder uitbreiden vanaf de plaats waar de lokale doorlaatbaarheidsverandering is opgetreden. De registratie-apparatuur geeft aan dat er kleine depolarisaties - ontladingen - van het membraan optreden. Deze worden exiterende postsynaptische potentialen (EPSP's) genoemd en lijken sprekend op de receptorpotentialen. Ze kunnen worden opgeteld en volgen niet de alles-of-niets-wet. [waardoor modulatie, regeling, mogelijk is]
Doordat de doorlaatbaarheid van het membraan alleen zeer lokaal verandert, treden er kleine stroompjes op die door de synaps lopen (de richting hiervan is aangegeven in figuur 47), en zich steeds verder uitbreiden vanaf de plaats waar de lokale doorlaatbaarheidsverandering is opgetreden. De registratie-apparatuur geeft aan dat er kleine depolarisaties - ontladingen - van het membraan optreden. Deze worden exiterende postsynaptische potentialen (EPSP's) genoemd en lijken sprekend op de receptorpotentialen. Ze kunnen worden opgeteld en volgen niet de alles-of-niets-wet. [waardoor modulatie, regeling, mogelijk is]Wanneer deze EPSP's boven een bepaalde drempelwaarde uitkomen, ontstaat in de motorische voorhoorncel een actiepotentiaal op de plaats waar de myelineschede van het axon begint [hilus]. De cel ontlaadt zich dus, hetgeen is aangegeven doordat cellichaam en axon rood getekend zijn en deze ontlading plant zich als een actiepotentiaal voort naar de spiervezels.
Wanneer treden actiepotentialen op in de motorische voorhoorncel? Dit kan op tweeërlei manier geschieden.
1. In de eerste plaats wanneer een lange reeks actiepotentialen in dezelfde afferente zenuwvezel aankomt en daardoor EPSP's veroorzaakt die opgeteld worden in het postsynaptische element, zodat het uiteindelijk effect voldoende is om de drempelwaarde te overschrijden. We spreken van temporale summatie omdat het een reeks prikkels betreft die over één enkele afferente zenuwvezel de motorische zenuwcel bereiken (de synaps is een 'ja-nee'-schakelaar).
2. In de tweede plaats, wanneer actiepotentialen over vele afferente zenuwvezels de motorische voorhoorncel bereiken, kan deze ook gedepolariseerd worden. We spreken dan van ruimtelijke summatie daar het prikkels betreft die over verschillende axonen bij de motorische cel aankomen. De excitatie is alleen dan effectief wanneer zij aanleiding geeft tot een actiepotentiaal in het postsynaptische element (de neuroncel is een optel-element). (64)
a. Inhibitie door een transmitterstof
Teneinde de inhibitie in de synaps te kunnen onderzoeken, moeten we gaan kijken naar de elektrische veranderingen in een van de motorische voorhoorncellen die bijdragen aan de innervatie van de strekkers van het bovenbeen. Wil de reflex gelijkmatig verlopen, dan moeten bij prikkeling van de voetzool bij de strandwandelaar deze spieren zich ontspannen.
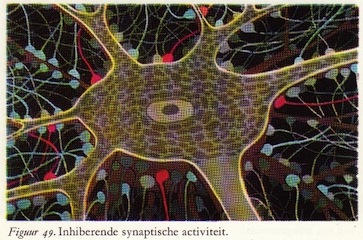
De actiepotentialen die aankomen in het presynaptische deel van de synaps - in dit geval de inhiberende of remmende synaps - zijn niet verschillend van die welke excitatie veroorzaakten.
Ter verduidelijking zijn de inhiberende eindigingen blauw aangegeven. Ook hier dus actiepotentialen die doodlopen in de eindvertakkingen van de presynaptische vezels. Het enige verschil is dat een andere soort transmitter- of overdrachtstof wordt vrijgemaakt: de inhiberende of remmende transmitterstof. Deze stof verandert ter plaatse de doorlaatbaarheid van het postsynaptische membraan en wel zo, dat speciaal de geleidbaarheid voor kalium- en chloorionen toeneemt. De ionenverschuivingen zijn dan ook tegengesteld aan die welke ogtreden bij excitatie, zoals door de blauwe en rode pijlen in fig. 53 en 54 is aangegeven. Ook hier zien we kleine synaptische stroompjes optreden die gaan lopen tussen de geactiveerde en de niet-geactiveerde delen van het postsynaptische membraan. (66)
Ter verduidelijking zijn de inhiberende eindigingen blauw aangegeven. Ook hier dus actiepotentialen die doodlopen in de eindvertakkingen van de presynaptische vezels. Het enige verschil is dat een andere soort transmitter- of overdrachtstof wordt vrijgemaakt: de inhiberende of remmende transmitterstof. Deze stof verandert ter plaatse de doorlaatbaarheid van het postsynaptische membraan en wel zo, dat speciaal de geleidbaarheid voor kalium- en chloorionen toeneemt. De ionenverschuivingen zijn dan ook tegengesteld aan die welke ogtreden bij excitatie, zoals door de blauwe en rode pijlen in fig. 53 en 54 is aangegeven. Ook hier zien we kleine synaptische stroompjes optreden die gaan lopen tussen de geactiveerde en de niet-geactiveerde delen van het postsynaptische membraan. (66)
b. Inhibitie door potentialen: hyperpolarisatie
De potentialen die we waarnemen door middel van de zeer dunne elektrode die zich in het cellichaam van de motorische voorhoorncel bevindt, zijn tegengesteld van teken als we ze vergelijken met de EPSP's. Ze worden inhiherende postsynaptische potentialen (IPSP's) genoemd.
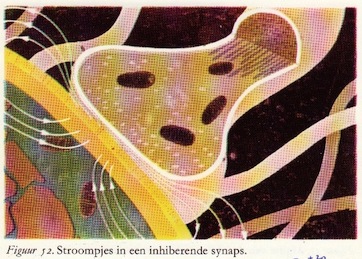
Daar de rustpotentiaal van het membraan negatief is (-60 mV) hebben ze tot gevolg dat deze membraanpotentiaal wordt verhoogd, meer negatief wordt, van -60 mV bijvoorbeeld tot -65 mV. Het membraan wordt dus niet ontladen, maar daarentegen meer negatief geladen: hyperpolarisatie. Dit verschijnsel kan dus nooit een actiepotentiaal opwekken, daar hiervoor het membraan juist gedepolariseerd of ontladen moet worden.
Het is nu gemakkelijk voor te stellen dat de kleine hyperpolarisaties even grote depolarisaties kunnen nivelleren. De totale som van de depolarisaties - de excitaties - moet ook altijd groter zijn dan de som van de hyperpolarisaties - de inhibities - teneinde actiepotentialen te kunnen opwekken in het postsynaptische neuron.
Het neuron zelf fungeert hier als een kleine telmachine. (68)
Het is nu gemakkelijk voor te stellen dat de kleine hyperpolarisaties even grote depolarisaties kunnen nivelleren. De totale som van de depolarisaties - de excitaties - moet ook altijd groter zijn dan de som van de hyperpolarisaties - de inhibities - teneinde actiepotentialen te kunnen opwekken in het postsynaptische neuron.
Het neuron zelf fungeert hier als een kleine telmachine. (68)
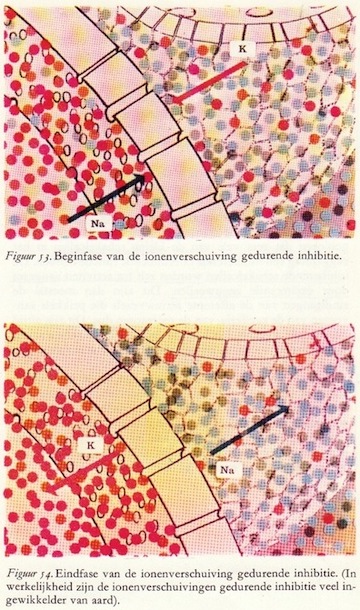
De zenuwcel: een microscopische rekenmachine
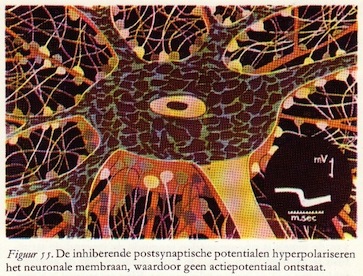 Vele axonen maken synaptisch contact met één enkele motorische voorhoorncel. Een deel van deze presynaptische vezels produceren exciterende potentialen, andere daarentegen inhiberende potentialen. De motorische zenuwcel telt de exciterende potentialen op, trekt daar de inhiberende vanaf en wanneer het totaal aanleiding geeft tot het overschrijden van de drempelwaarde, dan vuurt het axon van de motorische cel een of meerdere actiepotentialen of impulsen af naar de spier.
Vele axonen maken synaptisch contact met één enkele motorische voorhoorncel. Een deel van deze presynaptische vezels produceren exciterende potentialen, andere daarentegen inhiberende potentialen. De motorische zenuwcel telt de exciterende potentialen op, trekt daar de inhiberende vanaf en wanneer het totaal aanleiding geeft tot het overschrijden van de drempelwaarde, dan vuurt het axon van de motorische cel een of meerdere actiepotentialen of impulsen af naar de spier.De zenuwcellen en -vezels in de grijze stof van het ruggemerg zijn te verdelen in twee groepen, nl.
- de cellen die zorgen voor excitatie doordat ze exciterende transmitterstoffen afscheiden,
- en die welke zorgen voor inhibitie doordat ze remmende of inhiberende transmitterstoffen afscheiden.
Het is gebleken dat zeer vele van de kleine schakelcellen in het ruggemerg een inhiberende of remmende functie hebben. De inhiberende schakelcellen worden zelf tot activiteit aangezet door exciterende zenuwcellen. Dit zijn dan meestal de eindigingen van de afferente zenuwvezels, die prikkels aanvoeren van de periferie, van de receptoren dus. Dit zijn altijd exciterende prikkels. Het kunnen echter ook zenuwvezels zijn die ergens vanuit de hersenstam of vanuit de hersenschors hun invloed uitoefenen op de schakelcellen.
Samenvattend kunnen we zeggen dat in de synaptische activiteit van elke zenuwcel twee processen een belangrijke rol spelen: excitatie en inhibitie. Beide worden vergezeld van een zeer karakteristiek patroon van elektrische verschijnselen die tegengesteld aan elkaar gericht zijn. De basis waarop deze twee groepen verschijnselen zich afspelen, zijn transmitterstoffen die of exciterend of inhiberend werken. (70)
Prikkeloverdracht van zenuw op spier
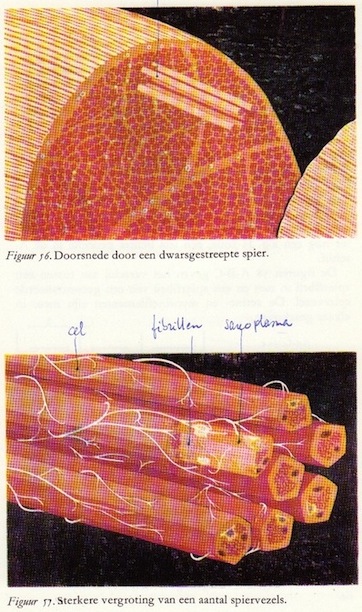 De prikkeloverdracht van de motorische of efferente zenuw op de spier en de daarop volgende contractie is alleen dan te begrijpen wanneer we ons eerst verdiepen in de macroscopische en microscopische bouw van de spier.
De prikkeloverdracht van de motorische of efferente zenuw op de spier en de daarop volgende contractie is alleen dan te begrijpen wanneer we ons eerst verdiepen in de macroscopische en microscopische bouw van de spier.Spieren bestaan uit naast elkaar liggende bundels spiervezels die door bindweefsel met elkaar verbonden zijn. Dikke bundels spiervezels worden omgeven door een zeer sterk bindweefselvlies; van hieruit gaan bindweefselschotten die kleinere bundels spiervezels omhullen, en tenslotte wordt elke spiervezel omgeven door zijn eigen bindweefselmembraan. (Opgebouwd zoals zenuwvezels)
In figuur 56 is een spier dwars doorgesneden. Een zevental spiervezels steekt uit het linkerdeel van de dwarsdoorsnede.
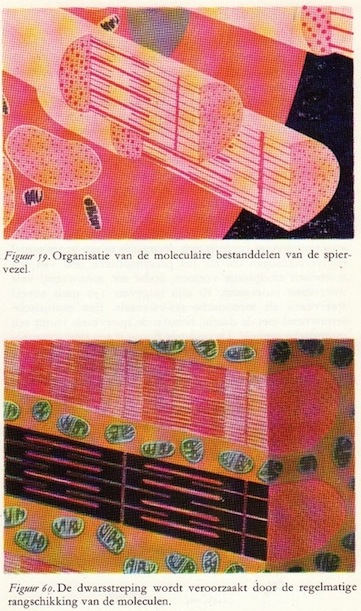
De onderste figuur geeft een sterkere vergroting hiervan: een van de vezels is opengesneden teneinde de inwendige structuur te kunnen laten zien. De witte draadjes zijn de eindvertakkingen van de motorische zenuw. Elke spiervezel is eigenlijk een langgerekte cilindrische cel die een lengte van 10-12 cm kan bereiken. De diameter is 10 tot l00μ. Elke vezel bevat verscheidene kernen die in het algemeen in de uiterste periferie van het sarcoplasma (het cytoplasma van de spiercel) gelegen zijn. Microscopisch blijkt de spiervezel een groot aantal dunne draadjes te bevatten: de spierfibrillen. Dit zijn de eigenlijke contractiele elementen van het spierweefsel. Deze dunne draadjes bleken - wanneer men gebruik maakte van de zeer sterke vergrotingen van het elektronenmicroscoop (meer dan 100.000x) te zijn opgebouwd uit filamenten, en wel:
a. dikke filamenten die bestaan uit het eiwit myoxine, en
b. dunne filamenten die bestaan uit het eiwit actine.
De actine- en de myosinefilamenten liggen keurig naast elkaar gerangschikt; ze zijn als het ware in elkaar geschoven. De dikke myosinefilamenten worden alle omgeven door zes dunne actinefilamenten. De lengte van een dergelijk actinefilament is ongeveer 1μ, terwijl die van een myosinefilament ongeveer 1,5μ bedraagt. (72)
Elektronenmicroscopie van de spiervezel
Het ultramicroscopische onderzoek van de spierfibril heeft nog een aantal andere eigenschappen aangetoond. In het midden van de actinefilamenten loopt dwars hierop het z.g.n. Z-membraan. De actinefilamenten lopen als het ware vanuit dit Z-membraan naar beide kanten tussen de myosinefilamenten in.
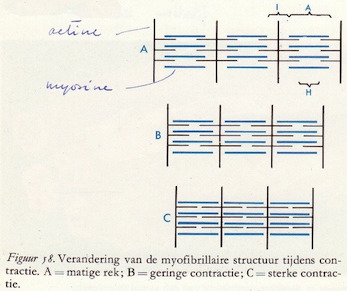
Men heeft dat deel van de spierfibril dat zich tussen twee Z-membranen in bevindt een sarcomeer genoemd.
We kunnen nu verder nog onderscheiden een A-schijf: dat deel van een sarcomeer waarin zich zowel actine- als myosinefilamenten bevinden, en verder een I-schijf waarin we alleen maar delen van de actinefilamenten tegenkomen. Het zijn nu deze structuurverschillen die de typische dwarsstreping aan de spieren geven. De A-schijven zien er onder de microscoop veel donkerder uit dan de I-schijven. Tevens onderscheiden we nog een z.g.n. H-zone, een iets lichter gedeelte in de A-schijf.
De figuren 58 A-B-C geven het verschil aan tussen een spierfibril in rust en een spierfibril van een gecontraheerde spiervezel. De actine- en myosinefilamenten zijn meer in elkaar geschoven. (74)
We kunnen nu verder nog onderscheiden een A-schijf: dat deel van een sarcomeer waarin zich zowel actine- als myosinefilamenten bevinden, en verder een I-schijf waarin we alleen maar delen van de actinefilamenten tegenkomen. Het zijn nu deze structuurverschillen die de typische dwarsstreping aan de spieren geven. De A-schijven zien er onder de microscoop veel donkerder uit dan de I-schijven. Tevens onderscheiden we nog een z.g.n. H-zone, een iets lichter gedeelte in de A-schijf.
De figuren 58 A-B-C geven het verschil aan tussen een spierfibril in rust en een spierfibril van een gecontraheerde spiervezel. De actine- en myosinefilamenten zijn meer in elkaar geschoven. (74)
De neuromusculaire synaps
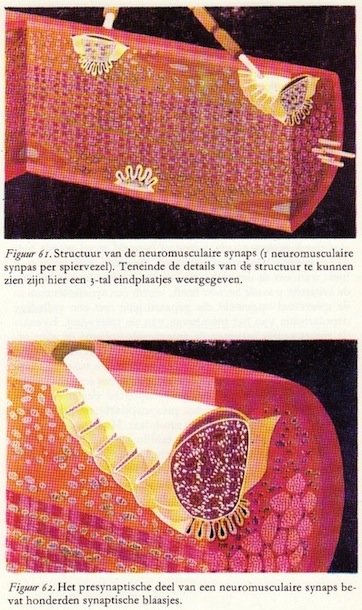 Nu wij iets meer te weten gekomen zijn omtrent de fijnere bouw van de spier gaan we een stap verder en bekijken de verbinding tussen de motorische zenuw en de spiervezels.
Nu wij iets meer te weten gekomen zijn omtrent de fijnere bouw van de spier gaan we een stap verder en bekijken de verbinding tussen de motorische zenuw en de spiervezels.De fijne uitlopers van de motorische zenuw maken contact met de spiervezels in de motorische eindplaat, ook wel neuromusculaire synaps of zenuwvezel/spiervezel-synaps genoemd. Deze wordt enerzijds gevormd door het trechtervormige, naakte uiteinde van de vertakkingen van de motorische zenuw (het presynaptische deel van de motorische eindplaat) en anderzijds door het in plooien liggende deel van het sarcolemma (het postsynaptische deel van de motorische eindplaat). Het sarcolemma is het membraan dat de spiervezel omgeeft. Deze bouw geeft de mogelijkheid van een zeer efficiënte prikkeloverdracht door het sterk vergrote oppervlak.
Een motorische zenuwvezel verdeelt zich doorgaans in een groot aantal eindvertakkingen die elk het presynaptische deel van een motorisch eindplaatje vormen, zodat één zenuwvezel vele spiervezels innerveert. Er zijn ongeveer 150 maal zoveel spiervezels als motorische zenuwvezels. Een motorische zenuwvezel met de daarbij behorende spiervezels vormt een motorische eenheid. De onderste figuur is een sterke vergroting van de bovenste. De meest opvallende structuurtjes zijn de kleine blaasjes die zich in het presynaptische deel van de motorische eindplaat bevinden. Het uiteinde van het axon is dus ook hier gevuld met de synaptische blaasjes die de transmitterstof bevatten, die ervoor zorgt dat de prikkel kan overspringen van het axon naar het postsynaptische deel van de spiervezel. De blaasjes zijn dezelfde die we al tegengekomen zijn in het presynaptische deel van de exciterende en inhiberende synaps. De chemische samenstelling is bij de neuromusculaire synaps geen geheim meer: de blaasjes bevatten acetylcholine. (76)
Spierpotentialen
 Wat de spiervezel betreft, heeft men ook kunnen vaststellen dat deze een rustpotentiaal bezit van 70-90 mV. Net als in de zenuwvezel is de binnenkant elektrisch negatief ten opzichte van de buitenkant. De rustpotentiaal is hier eveneens het gevolg van verschillen in ionenconcentraties tussen binnen- en buitenkant van het membraan van de spiervezel. Processen, analoog aan die in de zenuwvezel, zijn hiervoor verantwoordelijk.
Wat de spiervezel betreft, heeft men ook kunnen vaststellen dat deze een rustpotentiaal bezit van 70-90 mV. Net als in de zenuwvezel is de binnenkant elektrisch negatief ten opzichte van de buitenkant. De rustpotentiaal is hier eveneens het gevolg van verschillen in ionenconcentraties tussen binnen- en buitenkant van het membraan van de spiervezel. Processen, analoog aan die in de zenuwvezel, zijn hiervoor verantwoordelijk.Wordt de bijbehorende zenuw van een spier geprikkeld, dan nemen we in de motorische eindplaat een kleine verandering in de rustpotentiaal waar. Elke zenuwimpuls geeft een kleine depolarisatie of ontlading van het postsynaptische membraan. Dit is hetzelfde fenomeen dat we reeds tegenkwamen in de exciterende synaps. En wat dan met EPSP: exciterende postsynaptische potentiaal betiteld werd, wordt hier EPP: eindplaat-potentiaal genoemd. Dezelfde regel geldt ook hier: wanneer de depolarisatie van het membraan een bepaalde kritische waarde heeft bereikt, wordt een actiepotentiaal in de spiervezel opgewekt die gepaard gaat met een volledige depolariatie van het membraan van de spiervezel. Evenals de zenuwvezels reageren de spiervezels volgens de alles-of-niets-wet.
De EPP's geven aanleiding tot een actiepotentiaal die de spieractiepotentiaal wordt genoemd. De tussenschakel van de prikkel van de zenuw op de spier is het acetylcholine dat wordt vrijgemaakt van het presynaptische deel en terecht komt in de ruimte tussen het presynaptische en postsynaptische deel van de motorische eindplaat. Hierdoor wordt de ionendoorlaatbaarheid, de permeabiliteit, van het postsynaptische membraan veranderd. Natriumionen gaan naar binnen en kaliumionen treden uit de spiervezel. Hier speelt echter nog een ander ion een belangrijke rol, het calcium. Dit gedraagt zich op dezelfde wijze als het natrium: tijdens de spieractiepotentiaal gaat het van buiten naar binnen de zenuwvezel in. (78)
Van spierpotentiaal tot spiercontractie
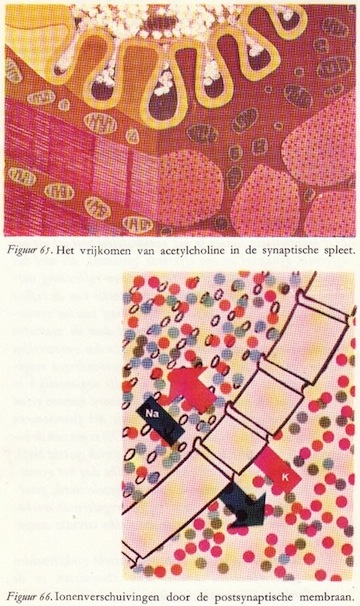 De spieractiepotentiaal is de vonk voor een chemische reactie in de spiervezel, waarbij het acetylcholine werkt als het ontstekingsmechanisme van de spiermotor. Kaliumionen treden uit de spiervezel en natrium- en calciumionen gaan naar binnen. De verhoging van de calciumionenconcentratie helpt mee aan het tot stand komen van een reactie waarbij de stof adenosinetrifosfaat (ATP) wordt gesplitst in adenosinedifosfaat (ADP) en fosforzuur. In symbolen uitgedrukt: ATP=ADP+P+E (E = energie); bij deze reactie komt energie vrij. De vrijgekomen energie wordt voornamelijk gebruikt om het actomyosinesysteem te verkorten, d.w.z.: de actinefilamenten schuiven verder tussen de myosinefilamenten in.
De spieractiepotentiaal is de vonk voor een chemische reactie in de spiervezel, waarbij het acetylcholine werkt als het ontstekingsmechanisme van de spiermotor. Kaliumionen treden uit de spiervezel en natrium- en calciumionen gaan naar binnen. De verhoging van de calciumionenconcentratie helpt mee aan het tot stand komen van een reactie waarbij de stof adenosinetrifosfaat (ATP) wordt gesplitst in adenosinedifosfaat (ADP) en fosforzuur. In symbolen uitgedrukt: ATP=ADP+P+E (E = energie); bij deze reactie komt energie vrij. De vrijgekomen energie wordt voornamelijk gebruikt om het actomyosinesysteem te verkorten, d.w.z.: de actinefilamenten schuiven verder tussen de myosinefilamenten in.Het ATP is de universele leverancier van energie in alle processen in alle cellen van het lichaam. Het wordt eerst in de mitochondria gevormd door de werking van de citroenzuurcyclus, waarin glucose en vetten worden omgezet in CO2 en water, en de daarbij vrijgekomen energie wordt vastgelegd in de fosforzuurverbinding ATP. Als bij een biochemische reactie energie nodig is, wordt ATP weer teruggevormd in ATP en fosforzuur, en de daarbij vrijkomende energie wordt benut om de reactie te laten verlopen.
De betekenis van de actiepotentiaal is in de spier een andere dan in de zenuw. In de zenuw brengt de actiepotentiaal een signaal over en is als het ware de functiedrager. ln de spiervezel echter gaat de functionele activiteit niet gepaard met signaal- of informatie-overbrenging, maar met contractie: de samentrekking van de spiervezel. De vrijgekomen chemische energie wordt door de spier omgezet in mechanische energie en de spier contraheert.
Er bestaan stoffen die de werking van acetylcholine remmen en deze stoffen, indien zij kunstmatig toegevoerd worden, remmen ook de prikkeloverdracht. Een ander feit is dat wanneer een microhoeveelheid acetylcholine op de motorische eindplaat wordt gedeponeerd, er een kleine depolarisatie optreedt, die identiek is aan de eindplaatpotentiaal. Wordt er een iets grotere hoeveelheid gedeponeerd, dan zien we de eindplaatpotentiaal toenemen en kan zelfs een spieractiepotentiaal worden opgewekt. Het blijkt nl. ook dat wanneer men lage concentraties acetylcholine gebruikt, de grootte van de depolarisatie evenredig is aan de concentratie. (80)
terug naar de Inhoud
3. Neuronale circuits
Neuronale plasticiteit
Het eenvoudigste circuit in het zenuwstelsel bestaat uit een afferente zenuwcel en een efferente zenuwcel, die met het ruggemerg als doorgangshuis tezamen een reflexboog vormen. Het inzicht in de structuur en functie van de reflex leverde een duidelijk beeld van de 'bedrading' van het zenuwstelsel. Deze bedrading wordt geleverd door de neurieten en dendrieten van de neuronen. De miljarden zenuwcellen maken een oneindig aantal onderlinge verbindingen mogelijk (die ook nog eens voortdurend veranderen: de neuronale plasticiteit; daarbij worden nieuwe dendrieten gevormd en ook weer afgebroken).
Alhoewel het spoor van vele circuits nog moeilijk te volgen is, zal een nadere analyse ons antwoord kunnen geven op een aantal vraagstukken betreffende het functioneren van ingewikkelde neuronale netwerken. Dit is een van de belangrijkste taken die de moderne hersenfysioloog zich gesteld heeft.
De miljarden bits aan informatie die elke dag het zenuwstelsel binnenkomen, moeten kritisch geselecteerd, gesorteerd en herinnerd worden, waarbij fijn regulerende mechanismen in het zenuwstelsel beslissen welke circuits aangesloten worden.
Een aantal neurologische en psychiatrische ziektebeelden kunnen worden geïnterpreteerd aan stoornissen in de 'bedrading' van neuronale netwerken. (82)
Functie van inhibitie in neuronale integratie
 Bij vertebraten vinden alle inhiberende processen plaats in het ruggemerg en in andere delen van het centrale zenuwstelsel. Wat de dwarsgestreepte spieren betreft, bestaat er geen perifere inhibitie maar uitsluitend centrale inhibitie.
Bij vertebraten vinden alle inhiberende processen plaats in het ruggemerg en in andere delen van het centrale zenuwstelsel. Wat de dwarsgestreepte spieren betreft, bestaat er geen perifere inhibitie maar uitsluitend centrale inhibitie.We moeten hierbij echter voor ogen houden dat, waar ook in het centrale en perifere zenuwstelsel, een neuron nooit minder dan niets kan doen. Ook de inhiberende mechanismen zijn dus actieve processen en deze worden op dezelfde wijze in gang gezet als de exciterende mechanismen, nl. door actiepotentialen van gelijke amplitude en gelijke duur, die in de presynaptische elementen opgewekt en voortgeleid worden. Inhibitie in het ruggemerg of andere delen van het zenuwstelsel kan het gevolg zijn van afferente prikkels uit de periferie, maar een even grote rol spelen hierbij de prikkels uit de verschillende delen van het centrale zenuwstelsel zelf.
De differentiatie in excitatie en inhibitie is een functie van de schakeling van de neuronen. Afhankelijk van het feit of ergens in de neuronenketen een inhiberende schakelcel geplaatst is, zal het uiteindelijke effect inhibitie zijn.
Inhibitie van een motore voorhoorncel kan geëffectueerd worden door verlaging van de prikkelbaarheid van die cel of door een reductie van de exciterende afferente signalen.
Wanneer echter een inhiberende schakelcel niet geactiveerd maar wordt geremd door een andere inhiberende schakelcel, dan kan de eerstgenoemde inhibitie uitgeschakeld worden, waardoor de motore voorhoorncel juist gefaciliteerd wordt; zo leidt 'dubbele inhibitie' tot een paradoxale reactie: facilitatie. De extra mogelijkheden die op deze manier aan het inhibitiemechanisme worden toegevoegd, kunnen alle op rekening geschreven worden van de specifieke eigenschappen van de schakelsystemen of neuronale circuits. (84)
De sturende taak van antidrome inhibitie
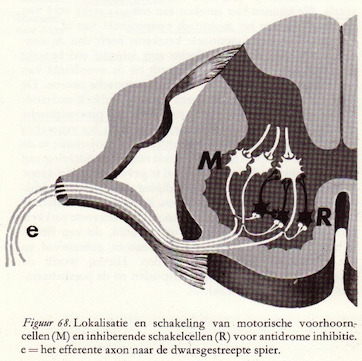 De structuur en functie van neuronale circuits is het best onderzocht in het ruggemerg, waar met behulp van microfysiologische methoden de inhiberende mechanismen geanalyseerd konden worden. De antidrome inhibitie is een voorbeeld van een eenvoudig, maar zeer vernuftig werkend systeem en een fraai voorbeeld van een cybernetisch regelsysteem. Ze is genoemd naar de Amerikaanse neurofysioloog Renshaw, die veel heeft bijgedragen aan onze kennis van de microfysiologie van het zenuwstelsel.
De structuur en functie van neuronale circuits is het best onderzocht in het ruggemerg, waar met behulp van microfysiologische methoden de inhiberende mechanismen geanalyseerd konden worden. De antidrome inhibitie is een voorbeeld van een eenvoudig, maar zeer vernuftig werkend systeem en een fraai voorbeeld van een cybernetisch regelsysteem. Ze is genoemd naar de Amerikaanse neurofysioloog Renshaw, die veel heeft bijgedragen aan onze kennis van de microfysiologie van het zenuwstelsel.De geëxciteerde motore voorhoorncellen sturen niet alleen actiepotentialen langs hun hoofdstam voor de stimulering van de dwarsgestreepte spieren, maar ook langs kleine collaterale zenuwvezels, die geheel binnen de grijze stof van het ruggemerg gelegen zijn. De eindvertakkingen van deze collateralen maken o.a. contact met de in de voorhoorn gelegen inhiberende schakelcellen, die op hun beurt de motore voorhoorncellen remmen. Wanneer de vuurfrequentie van de voorhoorncellen te hoog wordt, kan op deze manier de activiteit geremd worden.
Aan de uiteinden van de hoofdstam van het neuron (in de neuromusculaire synaps) en bij de prikkeloverdracht van de collateralen naar de inhiberende schakelcellen, wordt acetylcholine vrijgemaakt. De prikkeloverdracht van de inhiberende schakelcel naar de motore voorhoorncel vindt plaats door middel van een van de inhiberende neurotransmitters. Het remmende effect van deze schakelcellen duurt veel langer dan de gewone postsynaptische inhibitie; men heeft tijden gemeten van 50 tot 75 msec. Zowel schakeltechnisch als neurofysiologisch voldoet het systeem aan de grondprincipes van een cybernetisch model. Door negatieve feedback zorgt de teruggevoerde informatie dat een begaalde norm van de motore voorhoorncellen niet wordt overschreden. De signalen in het axonale of uitvoerende deel van het systeem controleren op deze manier de prikkelbaarheid van het gehele systeem, waardoor de vuurfrequentie van het axon altijd binnen nauwe (d.w.z. fysiologische) grenzen wordt gehouden. Bij uitschakeling van de antidrome inhibitie ontstaat dan ook een kramptoestand van de spier. (86)
Presynaptische inhibitie in een axo-axonale synaps
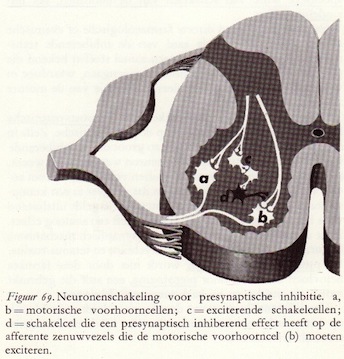 Een tweede voorbeeld van een remmend neuronencircuit is het presynaptische inhibitie-mechanisme, waarbij een axo-axonale synaps een grote rol speelt. Twee synapsen bevinden zich vlak bij elkaar, een axo-axonale en een axo-dendritische of axo-somatische. De actiepotentialen in de axo-axonale synaps verlagen de membraanpotentiaal (dit is dus een exciterend verschijnsel) van het presynaptische element van de axo-dendritische synaps.
Een tweede voorbeeld van een remmend neuronencircuit is het presynaptische inhibitie-mechanisme, waarbij een axo-axonale synaps een grote rol speelt. Twee synapsen bevinden zich vlak bij elkaar, een axo-axonale en een axo-dendritische of axo-somatische. De actiepotentialen in de axo-axonale synaps verlagen de membraanpotentiaal (dit is dus een exciterend verschijnsel) van het presynaptische element van de axo-dendritische synaps.De actiepotentialen die in dit presynaptische element arriveren, worden nu kleiner waardoor minder exciterende neurotransmitter-substantie wordt vrijgemaakt. Het depolariserende effect van dit tweede systeem wordt minder, waardoor uiteindelijk minder actiepotentialen door de motore voorhoorncel afgevuurd worden. Het postsynaptische neuron wordt dus niet direct door de presynaptische inhibitie beïnvloed, maar wel de signalen in de afferente vezels. We kunnen hier spreken van een selectieve remming van de input van het motorische systeem, hetgeen uiteraard een geheel andere functionele betekenis heeft dan de antidrome inhibitie, daar hier slechts zeer selectief een beperkt aantal afferente vezels geremd wordt bij de overdracht van hun exciterende invloed op het postsynaptische neuron.
De inhibitie wordt dus geëffectueerd door een dubbele excitatie: (met behulp van EPSP's) van een presynaptische eindiging (dit is dus een excitatie) vermindert het vrijmaken van een exciterende neurotransmitter. De depolarisatie in de axo-axonale synaps is juist voldoende om een verkleining van de amplitudo van de actiepotentiaal te geven, maar niet groot genoeg om de presynaptische eindiging van de axo-dendritische synaps geheel te depolariseren. Presynaptische inhibitie is vooral in het ruggemerg aangetoond. De afferente prikkels zijn voornamelijk pijnprikkels uit de huid, die een flexiereflex veroorzaken, waardoor alle flexoren geëxciteerd en alle extensoren geïnhibeerd worden. Hierbij wordt de regulerende invloed van de spierspoelen en de peeslichaampjes bijna geheel uitgeschakeld. (88)
Dubbele inhibitie veroorzaakt facilitatie
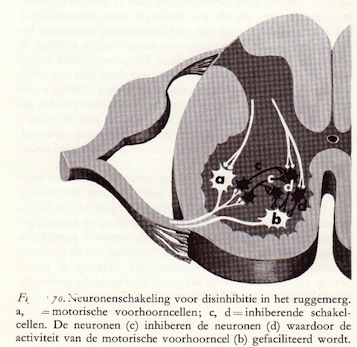 Zoals excitatie van een exciterend mechanisme inhibitie kan geven, zo is het ook mogelijk dat inhibitie van een inhiberend mechanisme resulteert in facilitatie. Hier spelen geen presynaptische maar uitsluitend postsynaptische verschijnselen een rol. Wanneer de inhiberende schakelcel van een inhiberend circuit door een andere inhiberende schakelcel, die deel kan uitmaken van een geheel ander circuit, wordt geremd, dan valt plotseling een deel van de inhibitie van de motore voorhoorncel weg, waardoor de exciterende prikkels een groter effect hebben, hetgeen een belangrijke facilitatie betekent. Vandaar de term disinhibitie, het vrijmaken van een inhiberend fenomeen, door inhibitie.
Zoals excitatie van een exciterend mechanisme inhibitie kan geven, zo is het ook mogelijk dat inhibitie van een inhiberend mechanisme resulteert in facilitatie. Hier spelen geen presynaptische maar uitsluitend postsynaptische verschijnselen een rol. Wanneer de inhiberende schakelcel van een inhiberend circuit door een andere inhiberende schakelcel, die deel kan uitmaken van een geheel ander circuit, wordt geremd, dan valt plotseling een deel van de inhibitie van de motore voorhoorncel weg, waardoor de exciterende prikkels een groter effect hebben, hetgeen een belangrijke facilitatie betekent. Vandaar de term disinhibitie, het vrijmaken van een inhiberend fenomeen, door inhibitie.Alhoewel de basisverschijnselen dus slechts tweeërlei zijn: excitatie en inhibitie, bepalen de neuronale circuits - de specifieke wijze van schakelen van de neuronen - het uiteindelijke effect.
Er zijn nu nog geen directe farmacologische of chemische aanwijzingen omtrent de aard van de inhiberende transmitter-substanties. Wel zijn er een aantal stoffen bekend die de post- of presynaptische inhibitie tegengaan, waardoor in vele gevallen een ongecontroleerde excitatie van de motore voorhoorncel het gevolg is.
De stof strychnine remt in belangrijke mate de postsynaptische inhibitie, maar heeft geen effect op de presynaptische. Zelfs in een geringe dosering is het effect zo groot, dat de inhiberende invloed van het postsynaptische neuron weggenomen wordt, zodat de normale exciterende impulsen voldoende zijn om zoveel actiepotentialen op te wekken, dat de spier in een kramptoestand geraakt. Dit effect van strychnine geldt uitsluitend voor het ruggemerg. Tetanus-toxine heeft een analoog effect.
Daar de antidrome inhibitie een postsynaptisch mechanisme is, wordt het ook beïnvloed door strychnine en tetanus-toxine.
De presynaptische inhibitie wordt niet door deze farmaca beïnvloed, maar wel door picrotoxine, een stof die gebruikt wordt als antidotum bij barbituraatvergiftigingen.
Het differentiële effect van deze farmaca duidt nadrukkelijk op het voorkomen van meer dan één inhiberende neurotransmitter-substantie. (90)
Schakelmogelijkheden voor 15 miljard eenheden
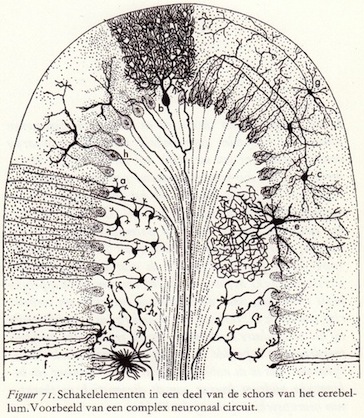 Leken de op de vorige bladzijden besproken neuronale circuits van het ruggemerg nog eenvoudig van aard, de systemen van de hersenen zelf, zoals de schors van de grote en kleine hersenen, zijn van een onmeetbare complexiteit. De miljarden neuronen in deze hersendelen hebben oneindig veel meer schakelmogelijkheden, doordat elke zenuwcel in synaptisch contact staat met tienduizenden andere neuronen.
Leken de op de vorige bladzijden besproken neuronale circuits van het ruggemerg nog eenvoudig van aard, de systemen van de hersenen zelf, zoals de schors van de grote en kleine hersenen, zijn van een onmeetbare complexiteit. De miljarden neuronen in deze hersendelen hebben oneindig veel meer schakelmogelijkheden, doordat elke zenuwcel in synaptisch contact staat met tienduizenden andere neuronen.De Engelse neuro-anatoom Young heeft de menselijke hersenen wel vergeleken met een kantoor met meer personeel dan er op dit moment aan mensen op de wereld rondlopen. Door middel van ongeveer 10 miljoen telefoonlijnen (vergelijkbaar met de 10 miljoen sensibele zenuwen die de grote hersenen bereiken) komt de informatie van de buitenwereld in codevorm het wereld-omvattende kantoor binnen. Hier bevinden zich dan 15 miljard typisten, secretaresses etc., die elk een zenuwcel vertegenwoordigen.
Op elke werktafel staat een telefooncentrale waardoor berichten van buiten het kantoor of van een andere afdeling de persoon in kwestie kunnen bereiken. Elk personeelslid brengt dan ook het grootste deel van zijn tijd door met het overbrengen van gecodeerde boodschappen naar collega's van zijn eigen afdeling of van andere afdelingen elders in het gebouw. Het grote aantal uitgaande lijnen dat met de buitenwereld verbonden is, zorgt, net als de motorische zenuwen, dat alle plannen, d.w.z. bewegingspatronen, stipt worden uitgevoerd.
Is het dan zo verwonderlijk dat, wanneer maar een klein aantal verbindingsdraden in deze zeer complexe 'warwinkel' van systemen defect raakt, de menselijke psyche uit evenwicht wordt gebracht? Temeer daar elke verbinding op zichzelf gevormd wordt door een minutieus geconstrueerde synaps, die op vele manieren te beïnvloeden is. Vandaar dat de moderne psycho-farmacologie zich voornamelijk richt op de bestudering van de synaptische activiteit en vooral de werkingswijze van de neurotransmitters. (92)
terug naar de Inhoud
4. Neurotransmitters
In de synaps vinden chemische en elektrische fenomenen plaats die bijzonder ingewikkeld verlopen en waarbij neurotransmitters een essentiële rol vervullen. De bestudering van het werkingsmechanisme van deze neurotransmitters is van groot belang voor het juiste begrip van de algemene werking van het zenuwstelsel.
De laatste jaren is het raakvlak met de klinische wetenschappen groter geworden, daar een aantal farmaca, zoals MAO-remmers, zowel bij de psychiater als bij de neurofysioloog, grote interesse hebben opgewekt. Er blijkt een gemeenschappelijke basis te bestaan daar deze farmaca enerzijds een verbetering van vitale depressies geven en anderzijds betrokken zijn bij het moleculaire mechanisme van de synaptische prikkeloverdracht in bepaalde delen van het zenuwstelsel. Mogelijkerwijs kan in de nabije toekomst de neurobioloog een belangrijke bijdrage leverenaan de diagnostiek en therapie van neuropsychiatrische ziektebeelden. (94)
De biochemische schakel in informatie-overdracht: de synaps
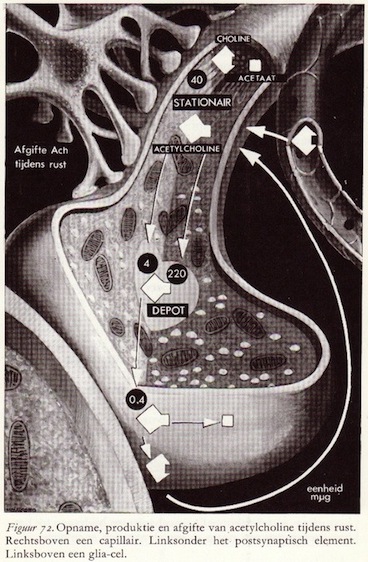 Alhoewel het mechanisme van het mobiliseren van de neurotransmitter-substantie door de actiepotentiaal in de presynaptische eindiging en de diffusie in de synapsspleet nog niet geheel tot klaarheid is gebracht, heeft men wel een indruk gekregen van de kwantitatieve verhoudingen die van betekenis zijn.
Alhoewel het mechanisme van het mobiliseren van de neurotransmitter-substantie door de actiepotentiaal in de presynaptische eindiging en de diffusie in de synapsspleet nog niet geheel tot klaarheid is gebracht, heeft men wel een indruk gekregen van de kwantitatieve verhoudingen die van betekenis zijn.Onder normale omstandigheden komt per minuut ongeveer 1-20 μg acetylcholine vrij in de synapsspleet van de exciterende synaps. Daar de duur en de amplitude van de actiepotentiaal even groot zijn, is er voldoende reden om aan te nemen dat elke actiepotentiaal een constante hoeveelheid neurotransmitter vrijmaakt. Voorlopige schattingen hebben uitgemaakt dat er zich tussen de 10.000 en 15.000 synaptische blaasjes bevinden in één enkele presynaptische eindiging. De presynaptische eindiging kan dus minuten lang werken zonder dat er nieuwe acetylcholine gesynthetiseerd behoeft te worden in het presynaptische element. De eenmaal vrijgekomen neurotransmitter-substantie wordt normaliter zeer snel afgebroken in de synapsspleet of op het postsynaptische membraan door middel van specitieke enzymwerkingen. De afgebroken bestanddelen zijn dan weer beschikbaar voor synthese van nieuwe neurotransmitter-substantie.
Wat betreft de aanwezigheid en het vrijkomen van neurotransmitter-substanties gelden nog de volgende regels:
1. Alle presynaptische eindigingen van een bepaald neuron bevatten dezelfde transmittersubstantie.
2. De neurotransmitters van twee achter elkaar geschakelde neuronen kunnen van elkaar verschillen in chemische samenstelling. (cholinergische, adrenergisch)
3. De eenmaal vrijgekomen neurotransmitter-substantie wordt normaliter snel afgebroken door enzymwerking. Deze enzymen (bij acetylcholine is dit het acetylcholine-esterase) zijn of aanwezig in het presynaptische, of in het postsynaptische element, of in de synaptische spleet.
4. De synaptische verbinding vormt een 'locus minoris resistentiae' (plaats met minder weerstand, het gevoelige punt) voor farmaca en toxinen, die op die plaats op meer of minder specifieke wijze kunnen aangrijpen. (96)
Analoog-digitaal-analoog-digitaal transmissie
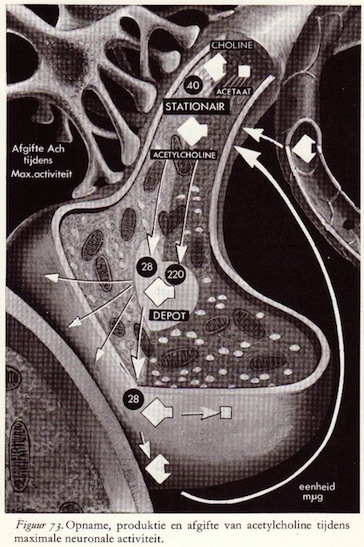 Wat betreft de chemische link in de synaptische transmissie zijn de volgende factoren nog van belang:
Wat betreft de chemische link in de synaptische transmissie zijn de volgende factoren nog van belang:- de chemische structuur van de neurotransmitter die aanwezig is in de presynaptische eindiging;
- de kwantiteit daarvan en de wijze waarop deze neurotransmitters kunnen worden vrijgemaakt uit de synaptische blaasjes en zodoende van de presynaptische eindiging kunnen diffunderen door de synaptische spleet naar het postsynaptische membraan;
- het aantal exciterende en inhiberende presynaptische eindigingen op het postsynaptische membraan en hun topografische organisatie.
De actiepotentiaal wordt dus in de synaps met behulp van de transmitterstof getransformeerd van een alles-of-nietssignaal in een in het postsynaptische element optredende elektrische verandering, die geheel andere eigenschappen heeft. Hier gebeurt dus het omgekeerde van wat bij de impulsoverdracht van zintuigcellen op afferente zenuwvezels plaatsvindt.
Om in rekenmachinetermen te spreken: het axon vertegenwoordigt het digitale type waarin informatie wordt weergegeven door elektrische impulsen. De synaps en het receptieve oppervlak van het postsynaptische element zijn dan te vergelijken met het analogetype waarin informatie wordt weergegeven door de sterkte van elektrische spanningen; de berekeningen worden uitgevoerd door het meten van die spanningen. Wij komen dus steeds de overgang analoog-digitaal-analoog tegen wanneer een impuls een neuronketen doorloopt.
Microchemisch onderzoek van de laatste jaren heeft een grote bijdrage geleverd tot de theoretische fundering dat de synaps niet meer beschouwd mag worden als een systeem met één-richtingsverkeer. De informatie-overdracht vindt weliswaar plaats van het presynaptische naar het postsynaptisehe element, maar op zijn beurt beïnvloedt het postsynaptische element ook de permeabiliteit en het potentiaalniveau van het presynaptische element. De synaps is dus een bijzondere vorm van een micro-cybernetisch systeem, waarbij tegenkoppeling- en terugkoppelingsmechanismen bijdragen tot het handhaven van een norm in een dynamisch evenwicht. (98)
Het autonome zenuwstelsel
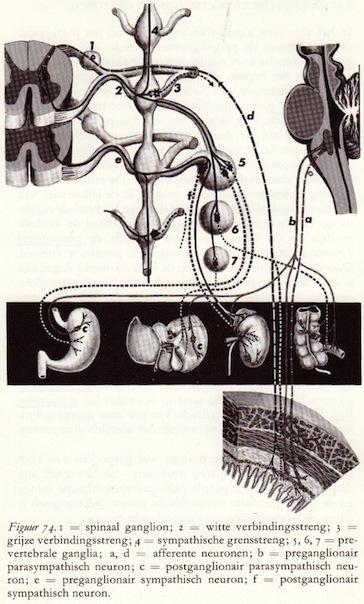 Het onderzoek naar andere neurotransmitters dan acetylcholine leidde reeds enkele decennia terug tot een gedetailleerd onderzoek van de structuur en functie van het autonome zenuwstelsel. Lang voordat men iets wist van de chemische samenstelling van transmitterstoffen was reeds bekend, dat vele autonome innervaties op twee - antagonistische - wijzen kunnen plaatsvinden, waarbij de neurohistoloog te hulp kwam en het gehele systeem in een sympathisch en een parasympathisch deel onderscheidde. Tot voor enkele jaren terug leek het alsof het autonome zenuwstelsel wat zijn structurele en functionele organisatie betreft aan enkele eenvoudige regels gehoorzaamde.
Het onderzoek naar andere neurotransmitters dan acetylcholine leidde reeds enkele decennia terug tot een gedetailleerd onderzoek van de structuur en functie van het autonome zenuwstelsel. Lang voordat men iets wist van de chemische samenstelling van transmitterstoffen was reeds bekend, dat vele autonome innervaties op twee - antagonistische - wijzen kunnen plaatsvinden, waarbij de neurohistoloog te hulp kwam en het gehele systeem in een sympathisch en een parasympathisch deel onderscheidde. Tot voor enkele jaren terug leek het alsof het autonome zenuwstelsel wat zijn structurele en functionele organisatie betreft aan enkele eenvoudige regels gehoorzaamde.Het zijn vooral een groep Zweedse onderzoekers geweest (Hillarp, Carlson en medewerkers) die met een nieuwe techniek, de fluorescentiemicroscopie, het klassieke beeld omver geworpen hebben. Zowel functie als structuur blijken veel complexer te zijn dan men aanvankelijk dacht. De belangrijkste ontdekking was wel dat niet alleen in het perifere autonome systeem nog een andere transmitter kon worden aangetoond (voor het sympathische systeem is dit adrenaline of noradrenaline) maar ook dat in alle delen van het centrale zenuwstelsel deze laatstgenoemde transmitter met een aantal andere nauwverwante stoffen een grote rol spelen bij de overdracht van signalen.
Voordat we meer in detail zullen ingaan op de nieuwe concepties betreffende de autonome prikkeloverdracht en de bespreking van de recentelijk ontdekte transmitters, zal een kort overzicht gegeven worden van de klassieke opvatting van de organisatie van het autonome zenuwstelsel, omdat dit zal dienen als basis voor de verdere bespreking.
De perifere delen van het sympathische en parasympathische zenuwstelsel bestaan uit twee neuronen die met elkaar synapteren in een perifeer ganglion. Het eerste neuron, dat naar het ganglion toeloopt, wordt de preganglionaire vezel genoemd. Het axon van het tweede neuron, waarvan het cellichaam ligt in het perifere ganglion, is de postganglionaire vezel. (100)
Cholinerge en adrenerge neuronen
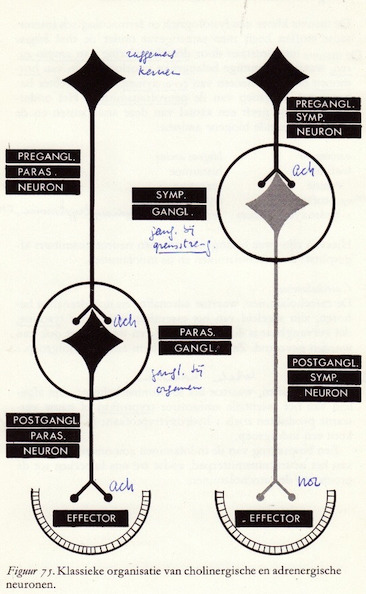 In het algemeen kunnen we stellen dat bij het parasympathische systeem de preganglionaire vezels lang zijn en de postganglionaire kort, omdat de ganglia altijd vlakbij of in de wand van het orgaan gelegen zijn. De belangrijkste parasympathische zenuw is de 10e hersenzenuw, de nervus vagus, maar er lopen ook parasympathische vezels mee met vele andere hersenzenuwen. Het sacrale deel van het parasympathische systeem bestaat uit dunne bundels zenuwvezels die ontspringen aan het 2e tot 4e sacrale ruggemergssegrnent.
In het algemeen kunnen we stellen dat bij het parasympathische systeem de preganglionaire vezels lang zijn en de postganglionaire kort, omdat de ganglia altijd vlakbij of in de wand van het orgaan gelegen zijn. De belangrijkste parasympathische zenuw is de 10e hersenzenuw, de nervus vagus, maar er lopen ook parasympathische vezels mee met vele andere hersenzenuwen. Het sacrale deel van het parasympathische systeem bestaat uit dunne bundels zenuwvezels die ontspringen aan het 2e tot 4e sacrale ruggemergssegrnent.Het sympathische deel van het autonome zenuwstelsel heeft een principieel andere organisatie. De cellichamen van de preganglionaire vezels zijn gelegen in de zijhoornen van het thoracale deel en het bovenste deel van het lumbale ruggemerg. De axonen van deze neuronen verlaten de ventrale wortels en de spinale zenuwen teneinde de grensstreng (truncus sympathicus) en de prevertebrale ganglia te vormen.
Daar de ganglia naast of voorop de wervelkolom gelegen zijn, is de verhouding pre- en postganglionaire vezels net andersom dan bij het parasympathische systeem: hier dus korte pre- en lange postganglionaire vezels.
Het verschil in functie tussen het sympathische en parasympathische zenuwstelsel wordt uitsluitend bepaald door het feit dat aan de uiteinden van de postganglionaire vezels verschillende neurotransmitters uitgescheiden worden. Voor het parasympathische systeem is dit acetylcholine, en voor het sympathische systeem voornamelijk noradrenaline; voor de prikkeloverdracht van pre- naar postganglionaire neuronen is in beide systemen het acetylcholine verantwoordelijk.
Kortheidshalve wordt daarom wel gesproken van cholinergische en adrenergische neuronen - cholinergisch zijn dan de pre- en de postganglionaire parasympathische zenuw en de preganglionaire sympathische zenuw; adrenergisch is alleen de postganglionaire sympathische zenuw. (102)
Biogene aminen als neurotransmitters
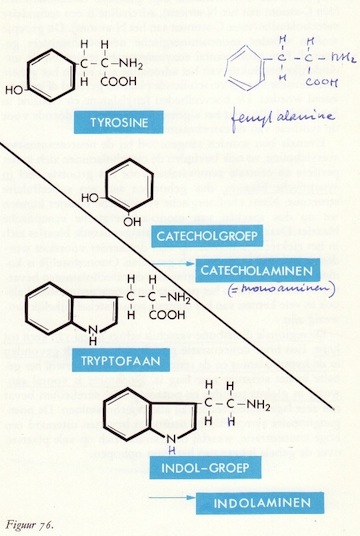 De nieuwe klasse van fysiologisch en farmacologisch interessante stoffen heeft men samengevat onder de titel biogene aminen. Deze ontstaan door decarboxylering van amino-zuren, waarvan sommige belangrijke voorlopers zijn van hormonen of componenten van co-enzymen; andere echter behoren tot de groep van de neurotransmitters. Het onderstaande lijstje geeft een aantal van deze aminozuren en de daarvan afgeleide biogene aminen:
De nieuwe klasse van fysiologisch en farmacologisch interessante stoffen heeft men samengevat onder de titel biogene aminen. Deze ontstaan door decarboxylering van amino-zuren, waarvan sommige belangrijke voorlopers zijn van hormonen of componenten van co-enzymen; andere echter behoren tot de groep van de neurotransmitters. Het onderstaande lijstje geeft een aantal van deze aminozuren en de daarvan afgeleide biogene aminen:aminozuur biogene amine
histidine histamine
tyrosine tyramine
tryptofaan tryptamine
5-hydroxytryptofaan serotonine
Hiervan zijn twee belangrijke groepen neurotransmitters afgesplitst: de catecholaminen en de indolaminen.
1. Catecholaminen
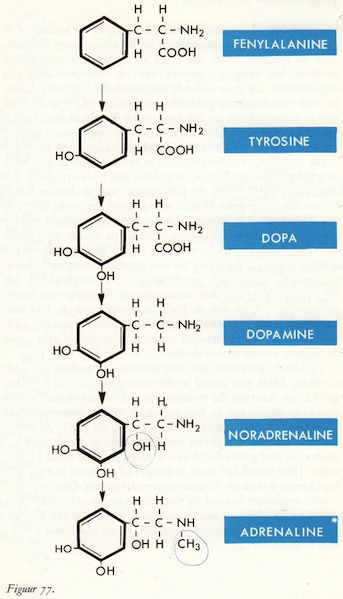 De catecholaminen, waartoe adrenaline en noradrenaline behoren, zijn afgeleid van het essentiele aminozuur tyrosine, dat vervangbaar is door fenylalanine en waaruit het ook kan worden gevormd. Ze bevatten als kern een catecholgroep.
De catecholaminen, waartoe adrenaline en noradrenaline behoren, zijn afgeleid van het essentiele aminozuur tyrosine, dat vervangbaar is door fenylalanine en waaruit het ook kan worden gevormd. Ze bevatten als kern een catecholgroep.2. Indoaminen
De indolaminen, waartoe het serotonine behoort, zijn afgeleid van het essentiële aminozuur tryptofaan of nauw verwante producten zoals 5-hydroxytryptofaan; ze bevatten als kern een indolgroep.
Een bespreking van de indolaminen zou ons te ver voeren van het neurotransmitterpad, zodat we ons beperken tot de groep van de catecholaminen. (104)
Catecholaminen
Het nevenstaande schema laat de vorming zien van de catecholaminen uit de aminozuren fenylalanine en tyrosine. Dopamine en noradrenaline zijn primaire catecholaminen (één C-atoom aan het N-atoom), adrenaline is een secundaire catecholamine (twee C-atomen aan het N-atoom). Dit groepje stoffen worden monoaminergische neurotransmitters genoemd. Een groot aantal enzymsystemen spelen bij de uiteindelijke productie van het adrenaline een rol en het gehele mechanisme kan op verschillende plaatsen geremd of geblokkeerd worden. De hoeveelheden fenylalanine en tyrosine in het voedsel zijn over het algemeen meer dan voldoende voor de synthese van de catecholaminen.
Evenals kon worden aangetoond bij de neurotransmitter acetylcholine, zo ook bevinden de catecholaminen zich in het perifere en centrale zenuwstelsel voor het grootste deel in synapstische blaasjes, dus gebonden aan een subcellulaire structuur. Naast cholinergische synaptische blaasjes kunnen we nu dus spreken van mono-aminergische synaptische blaasjes. Daar de inhoud van de laatstgenoemde blaasjes zich in het elektronenmicroscoop veel donkerder voordoet, worden ze ook wel aangeduid als granulae.
Onomstotelijk is vast komen te staan dat deze granulae de catecholaminen bevatten en dat ze zowel in het perifere autonome zenuwstelsel alsook in vele kernen van het centrale zenuwstelsel rijkelijk aanwezig zijn. De regionale distributie verschilt echter nogal van kern tot kern. Een hoge concentratie noradrenaline wordt gevonden in de hypothalamus en de reticulaire formatie, terwijl het gehalte in het striatum zeer laag is. Dopamine is vooral aanwezig in globus pallidus en putamen. Het cerebellum bevat een zeer lage concentratie van alle catecholaminen. De postganglionaire sympathische neuronen bevatten uiteraard een hoge concentratie, waarbij de granulae zich op vele plaatsen over de gehele lengte van het axon ophopen. (106)
Fenylalanine → tyrosine
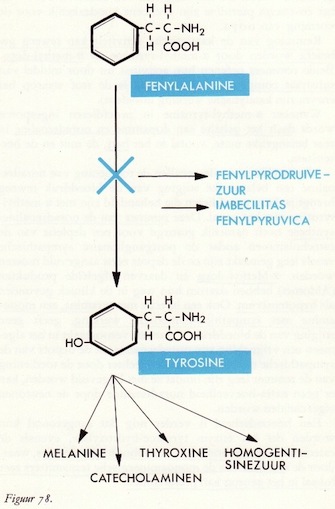 De bestudering van de afzonderlijke stappen die leiden tot de vorming van de catecholaminen uit de aminozuren fenylalanine en tyrosine heeft een schat van gegevens opgeleverd omtrent het aangrijpingspunt van de monoaminergische transmitters. De verschillende enzymatische stappen kunnen op zeer vele manieren geblokkeerd worden en het is dan ook niet te verwonderen dat het speurwerk naar deze reacties een aantal belangrijke geneesmiddelen heeft opgeleverd en ook onze kennis omtrent een aantal ziektebeelden heeft verdiept.
De bestudering van de afzonderlijke stappen die leiden tot de vorming van de catecholaminen uit de aminozuren fenylalanine en tyrosine heeft een schat van gegevens opgeleverd omtrent het aangrijpingspunt van de monoaminergische transmitters. De verschillende enzymatische stappen kunnen op zeer vele manieren geblokkeerd worden en het is dan ook niet te verwonderen dat het speurwerk naar deze reacties een aantal belangrijke geneesmiddelen heeft opgeleverd en ook onze kennis omtrent een aantal ziektebeelden heeft verdiept.De overgang van fenylalanine in tyrosine wordt gekatalyseerd door een oxygenase, waardoor een hydroxylgroep geintroduceerd wordt in de parapositie van de benzeenring. Indien deze stap geblokkeerd wordt door het ontbreken van het desbetreffende enzym, dan ontstaat het tussenproduct fenylpyrodruivezuur, hetgeen in de urine wordt uitgescheiden. Deze pathologische conditie wordt fenylketonurie genoemd en geeft aanleiding tot een ernstige vorm van krankzinnigheid (imbicilitas fenylperuvica). Afwezigheid in het voedsel van fenylalanine voorkomt de vorming van fenylpyrodruivezuur en ook de pathologische conditie. Het ontbreken van het genoemde enzym is genetisch bepaald, vandaar dat men dit ziektebeeld rangschikt onder de groep van de 'inborn errors of metabolism'.
Tyrosine, gevormd uit fenylalanine of rechtstreeks uit eiwitten, heeft een aantal stofwisselingswegen tot zijn beschikking, waarvan de vorming van de catecholaminen (over DOPA) er slechts één is. Een weg leidt over het tussenproduct homogentisinezuur uiteindelijk tot CO2 en H2O. Alkaptonurie is een pathologische conditie, waarbij het homogentisinezuur niet geoxydeerd kan worden en in de urine terecht komt. Hier wordt het zuur na enige tijd omgezet in een donker chinon, waardoor de urine inktzwart wordt. Daar tyrosine verder essentieel is voor de vorming van het schildklierhormoon en het pigment melamine, zijn de gevolgen van het ontbreken van tyrosine (cretinisme, albinisme) merkbaar in zeer veel orgaanfuncties. (108)
Tyrosine → DOPA
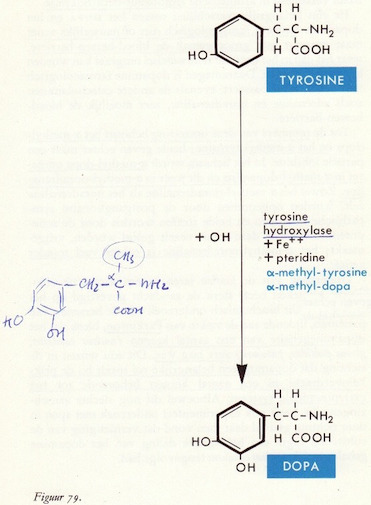 Het enzym tyrosone-hydroxylase katalyseert de vorming van 3.4-dihydroxyfenylalanine (afgekort tot DOPA); er komt nu een tweede OH-groep bij in de benzeenring. Deze overgang bepaalt tevens de snelheid waarmee uiteindelijk noradrenaline wordt gevormd. De aanwezigheid van zuurstof, Fe++ en het co-enzym pteridine zijn eveneens noodzakelijk voor de vorming van DOPA.
Het enzym tyrosone-hydroxylase katalyseert de vorming van 3.4-dihydroxyfenylalanine (afgekort tot DOPA); er komt nu een tweede OH-groep bij in de benzeenring. Deze overgang bepaalt tevens de snelheid waarmee uiteindelijk noradrenaline wordt gevormd. De aanwezigheid van zuurstof, Fe++ en het co-enzym pteridine zijn eveneens noodzakelijk voor de vorming van DOPA.Remming van de katalytische activiteit kan teweeggebracht worden door alfa-methyl-tyrosine en alfa-methyl-dopa. Beide remmers oefenen hun activiteit uit door middel van substraat competitie (het substraat is de stof waarop het enzym zijn katalytische werking uitoefent). Wanneer alfa-methyl-tyrosine in proefdieren wordt ingespoten, daalt het gehalte aan dopamine en noradrenaline in belangrijke mate, vooral in het hart, de milt en de hersenstam.
Alhoewel in normale gevallen de toediening van noradrenaline een belangrijke stijging van de bloeddruk teweeg brengt, is dit in proefdieren die behandeld zijn met alfa-methyl-tyrosine niet het geval. Deze remmer van de noradrenalinesynthese heeft namelijk gezorgd voor een depletie van de catecholaminen, zodat de postganglionaire sympathische vezels leeg gemaakt zijn en de depots eerst aangevuld moeten worden. alfa-Methyl-dopa en daarvan afgeleide producten hebben daarom hun weg in de kliniek gevonden als hypotensivum. Ook een injectie met tyramine, mono-amine met sympathico-mimetische werking, geeft geen stijging van de bloeddruk. Tyramine veroorzaakt in het algemeen een vrijmaking van noradrenaline uit de depots van de sympathische neuronen; daar deze echter door de toediening van de remmer leeg zijn doordat ze niet worden bijgevuld, kan er geen extra-hoeveelheid noradrenaline door de neuronen afgescheiden worden.
Een bijzonderheid is verder nog dat kon worden aangetoond dat het enzym tyrosine-hydroxylase, evenals de catecholaminen, gebonden is aan subcellulaire deeltjes, waardoor de synthese van de monoaminergische transmitters zeer lokaal in het neuron kan plaatsvinden. (110)
DOPA → dopamine
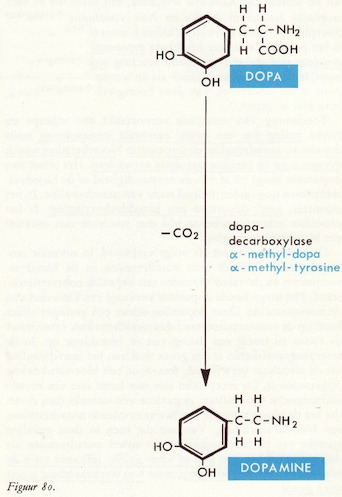 De omzetting van DOPA in dopamine wordt gekatalyseerd door dopa-decarboxylase. Daar dit enzym vele andere derivaten van de catecholaminen en eveneens 5-hydroxytryptofaan tot serotonine kan omzetten, heeft men recentelijk de naam veranderd in aromatische aminozuur-decarboxylase.
De omzetting van DOPA in dopamine wordt gekatalyseerd door dopa-decarboxylase. Daar dit enzym vele andere derivaten van de catecholaminen en eveneens 5-hydroxytryptofaan tot serotonine kan omzetten, heeft men recentelijk de naam veranderd in aromatische aminozuur-decarboxylase.Er zijn belangrijke verschillen tussen het DOPA en het dopamine. DOPA is farmacologisch niet of nauwelijks actief, maar passeert met groot gemak de bloed-hersen-barrière, waar het dan in het centrale zenuwstelsel omgezet kan worden in catecholaminen. Daarentegen is dopamine farmacologisch zeer actief, maar passeert, evenals de andere catecholaminen zoals adrenaline en noradrenaline, zeer moeilijk de bloed-hersen-barriere.
Tot de remmers van deze omzetting behoort het alfa-methyl-dopa en het alfa-methyl-tyrosine; beide geven echter maar een partiële inhibitie. In het lichaam wordt alfa-methyl-dopa omgezet in alfa-methyl-dopamine en dit weer in alfa-methyl-noradrenaline. Zowel het alfa-methyl-noradrenaline als het noradrenaline zelf, worden opgenomen door de postganglionaire sympathische neuronen en beide stoffen worden door de actiepotentialen die in deze zenuwvezels geleid worden, vrijgemaakt; het alfa-methyl-noradrenaline is echter veel minder actief.
Onderzoek van de laatste jaren in een aantal neurologische klinieken heeft sterk de aandacht gevestigd op het dopamine. Uit biochemisch onderzoek van de hersenen van patiënten, lijdende aan de ziekte van Parkinson, bleek dat het dopaminegehalte van een aantal kernen (nucleus caudatus, globus pallidus, putamen) zeer laag was. Dit zou wijzen in de richting dat dopamine een belangrijke rol speelt bij de prikkeloverdracht in een aantal kernen, behorende tot het extrapiramidale systeem. Alhoewel dit nog slechts aanwijzingen zijn, heeft ook experimenteel onderzoek met apen in deze richting geduid, daar men vond dat vernietiging van de substantia nigra een belangrijke daling van het dopamine gehalte van de nucleus caudatus tengevolge had. (112)
Dopamine en de perifere circulatie
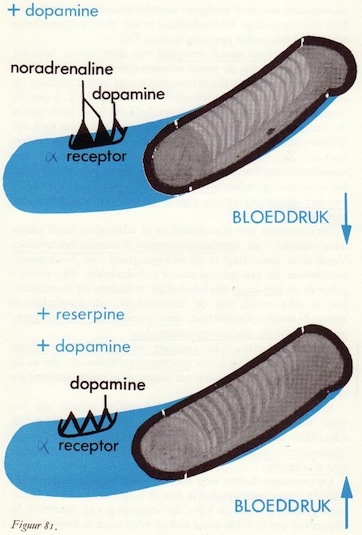 Een injectie met dopamine heeft een dualistisch effect op het vasculaire apparaat, afhankelijk van het feit of het proefdier voorbehandeld is met reserpine. Geen farmacon heeft ons inzicht in de problematiek van de werkingssfeer van de catecholaminen zo verdiept als reserpine. Het is een alkaloïd uit de wortel van Rauwolfia Serpentina, een plant die in vele tropische landen in Afrika en Azië voorkomt. Alhoewel reserpine nauwelijks klinisch meer wordt toegepast, is het gebleken een bruikbaar hypertensivum en neurolepticum (een stof die een opvallend krachtig sedatief vermogen bezit) te zijn. Zowel uit klinisch als uit wetenschappelijk oogpunt is het onderzoek van deze stof nog lang niet uitgeput.
Een injectie met dopamine heeft een dualistisch effect op het vasculaire apparaat, afhankelijk van het feit of het proefdier voorbehandeld is met reserpine. Geen farmacon heeft ons inzicht in de problematiek van de werkingssfeer van de catecholaminen zo verdiept als reserpine. Het is een alkaloïd uit de wortel van Rauwolfia Serpentina, een plant die in vele tropische landen in Afrika en Azië voorkomt. Alhoewel reserpine nauwelijks klinisch meer wordt toegepast, is het gebleken een bruikbaar hypertensivum en neurolepticum (een stof die een opvallend krachtig sedatief vermogen bezit) te zijn. Zowel uit klinisch als uit wetenschappelijk oogpunt is het onderzoek van deze stof nog lang niet uitgeput.Toediening van reserpine veroorzaakt een sterke daling van een aantal cerebrale monoaminen zoals dopamine, noradrenaline en serotonine. Noradrenaline wordt eveneens uit de perifere circulatie verwijderd. Het effect van dopamine hangt nu af van de omstandigheid of de bloedvatreceptoren nog onder invloed staan van noradrenaline. In het algemeen geeft dopamine een bloeddrukverlaging. Is het proefdier echter voorbehandeld met reserpine dan ontstaat een bloeddrukverhoging.
Dit fenomeen wordt als volgt verklaard. In normale omstandigheden bevindt zich noradrenaline in de bloedvatreceptoren en de vaten vertonen een bepaalde contractietoestand. Het toegediende dopamine vervangt een klein deel van de noradrenaline. Daar dopamine echter een geringer effect heeft op de contractietoestand dan noradrenaline, vermindert de tonus en treedt een daling van de bloeddruk op. In de reserpineproefdieren is een groot deel van het noradrenaline uit de circulatie verwijderd, waardoor een bloeddrukdaling opgetreden is. De receptoren zijn niet bezet met een monoaminergische transmitter; dopamine veroorzaakt dan, doordat het de plaats inneemt van het verwijderde noradrenaline, een bloeddrukstijging. Vandaar dat men in deze gevallen spreekt van partiële antagonisten: zowel noradrenaline als dopamine hebben een min of meer gelijke affiniteit voor de receptoren van de bloedvaten, maar hun werkzaamheid is niet even groot. (114)
Dopamine → noradrenaline → adrenaline
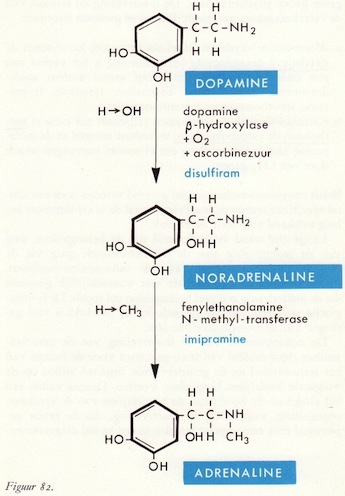 De omzetting van dopamine in noradrenaline gebeurt met behulp van dopamine-ß-hydroxylase. Ook dit enzym is gebonden aan subcellulaire deeltjes. Voor de omzetting is moleculaire zuurstof nodig en ascorbinezuur is een cofactor.
De omzetting van dopamine in noradrenaline gebeurt met behulp van dopamine-ß-hydroxylase. Ook dit enzym is gebonden aan subcellulaire deeltjes. Voor de omzetting is moleculaire zuurstof nodig en ascorbinezuur is een cofactor.Een merkwaardige bijzonderheid is dat deze enzymatische omzetting bijzonder gevoelig is voor CO.
Er is een groot aantal remmers van deze reactie bekend, waarvan disulfiram de meest interessante is. Worden proefdieren behandeld met disulfiram, dan hoopt zich dopamine op en het gehalte aan noradrenaline wordt zeer laag. Voor de studie van het catecholaminenmetabolisme in de hersenen is dit van groot belang, omdat disulfiram ons in staat stelt het differentiële effect van het verschil in concentraties te onderzoeken. Zo verlaagt bijvoorbeeld alfa-methyl-tyrosine het noradrenalinegehalte in de hersenen, maar beïnvloedt niet het serotoninegehalte; disulfiram veroorzaakt een verhoginig van het dopamine- en een verlaging van het noradrenaline gehalte.
De conversie van noradrenaline in adrenaline heeft plaats door middel van fenylethanolamine-N -methyl-transferase. Voor deze omzetting is de aanwezigheid van S-adenosylmethionine als een methyl-donor noodzakelijk. Het vermogen van de bijnieren om adrenaline te maken uit noradrenaline is afhankelijk van de concentratie van fenylethanolamine-N-methyl-transferase; deze concentratie wordt gereguleerd door de glucocorticosteroïden. De verhouding noradrenaline-adrenaline wisselt van orgaan tot orgaan: het bijniermerg scheidt voornamelijk adrenaline af met zeer weinig noradrenaline, terwijl de sympathische postganglionaire neuronen vooral noradrenaline afscheiden.
De bovengeschetste weg voor de synthese van de catecholaminen in het zenuwstelsel is niet de enige maar wel de meest belangrijke; zo schijnt bijv. de omzetting van tyramine in dopamine wel in vitro maar niet in vivo voor te komen. (116)
De afbraak van catecholaminen
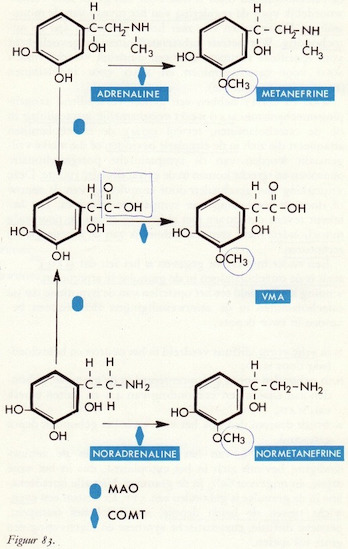 De afbraak van de catecholaminen staat, misschien nog meer dan de synthese, reeds jaren in het middelpunt van de belangstelling omdat de ontdekking van een aantal specifieke remmers van de enzymen die voor deze afbraak verantwoordelijk zijn, aanleiding gegeven heeft tot de ontwikkeling van een grote groep psychofarmaca. De inactivering en afbraak van de catecholaminen geschiedt door twee groepen enzymen:
De afbraak van de catecholaminen staat, misschien nog meer dan de synthese, reeds jaren in het middelpunt van de belangstelling omdat de ontdekking van een aantal specifieke remmers van de enzymen die voor deze afbraak verantwoordelijk zijn, aanleiding gegeven heeft tot de ontwikkeling van een grote groep psychofarmaca. De inactivering en afbraak van de catecholaminen geschiedt door twee groepen enzymen:a. Mono-amine-oxydasen (afgekort tot MAO) katalyseren de oxydatieve desaminering (desaminering is het verlies van een aminogroep) van een groot aantal stoffen zoals: dopamine, noradrenaline, adrenaline, tyramine, tryptamine, serotonine en andere monoaminen.
b. Catechol-O-methyl-transferasen (afgekort tot COMT) methyleren de catecholaminen, waardoor meestal in de metapositie van de benzeenring een H-atoom vervangen wordt door een CH3-groep.
Beide enzym-systemen kunnen geremd worden door een aantal specifieke remmers, waarvan vooral de MAO-remmers van belang zijn voor de kliniek.
Lange tijd werd verondersteld dat de belangrijkste weg van de inactivering van de catecholaminen ging door de desaminering met behulp van de monoamine-oxydasen. Recent onderzoek heeft echter zeer waarschijnlijk gemaakt dat de methylering een veel belangrijker rol speelt. De fysiologische activiteit van het gemethyleerde product is veel geringer dan de oorspronkelijke stof.
De consequenties van de inactivering van de catecholaminen door middel van MAO en COMT voor de functie van het zenuwstelsel en de geïnnerveerde organen, zullen op de volgende bladzijden besproken worden. Daarna zullen aan het eind van dit hoofdstuk de hoofdlijnen van de synthese, opstapeling, vrijmaking en inactivering, die de revue gepasseerd zijn, samengevat worden in een aantal diagrammen. (118)
Monoamine-oxydasen (MAO)
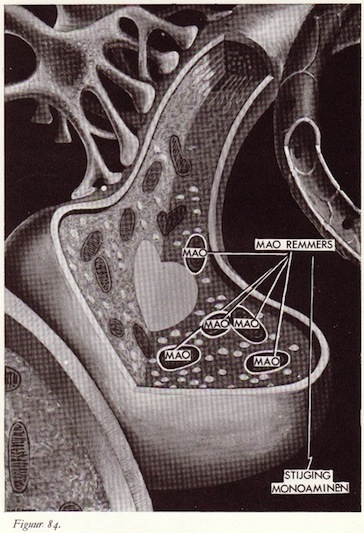 MAO katalyseert niet alleen de oxydatieve desaminering van de catecholaminen, maar is ook voor een groot deel verantwoordelijk voor de regulering van het niveau van de monoaminen. De enzymen zijn gebonden aan de mitochondria. Wat betreft substraataffiniteit en gevoeligheid voor specifieke remmers is er een duidelijk verschil tussen MAO voor catecholaminen en MAO voor indolaminen (tryptamine, serotonine).
MAO katalyseert niet alleen de oxydatieve desaminering van de catecholaminen, maar is ook voor een groot deel verantwoordelijk voor de regulering van het niveau van de monoaminen. De enzymen zijn gebonden aan de mitochondria. Wat betreft substraataffiniteit en gevoeligheid voor specifieke remmers is er een duidelijk verschil tussen MAO voor catecholaminen en MAO voor indolaminen (tryptamine, serotonine).MAO en COMT hebben een geheel verschillend aangrijpingsmechanisme: MAO werkt voornamelijk intracellulair in op de catecholaminen, terwijl COMT inwerkt op de catecholaminen die zich in de circulatie bevinden of welke vrijgemaakt worden van de sympathische postganglionaire neuronen en terecht komen in de extracellulaire ruimte. Deze vrijmaking kan geschieden door stimulering van de zenuw of door de toediening van sympathico-mimetica. Dit betekent tevens dat COMT niet gebonden is aan de neuronale mitochondria, maar zeer waarschijnlijk aan de adrenerge receptoren.
Een ander interessant gegeven is het feit dat MAO niet in staat is de catecholaminen in de granulae om te zetten. Deze vinding heeft geleid tot het opstellen van de hypothese dat catecholaminen in de zenuweinden zich kunnen bevinden in twee depots:
a. in vrije vorm, diffuus verdeeld in het neuron en beïnvloedbaar door MAO;
b. in gebonden vorm, geconcentreerd in de granulae, gebonden aan ATP in een verhouding van 4:1 en buiten bereik van MAO;
a. wordt daarom dan ook het vrije en b. het gebonden depot genoemd.
Ongeveer 40% van het noradrenaline in de zenuweinden bevindt zich in het cytoplasma, dus in het vrije depot, en ongeveer 60% in de granulae. Niet alle noradrenaline in de granulae is gebonden aan ATP. Er bestaat een evenwicht tussen de beide depots, waarbij actief transport, passieve diffusie, enzymatische synthese en inactivering een grote rol spelen. (120)
MAO remmers
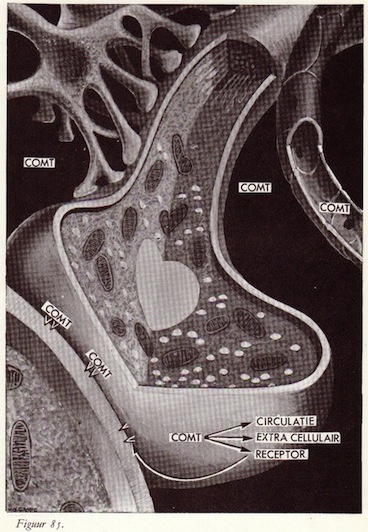 Het ligt voor de hand dat van de remmers van MAO verwacht mag worden dat zij een grote invloed hebben op de stofwisseling van de monoaminen. Na toediening van MAO-remmers bij proefdieren is dan ook gebleken dat de concentratie aan monoaminen in het centrale zenuwstelsel sterk stijgt; verscheidene honderden MAO-remmers zijn in de laatste jaren door farmaceutische laboratoria gesynthetiseerd, een klein percentage ervan is van blijvende waarde gebleken. De meeste thans in gebruik zijnde MAO-remmers zijn hydrazinederivaten die worden gekenmerkt door de hydrazinegroep.
Het ligt voor de hand dat van de remmers van MAO verwacht mag worden dat zij een grote invloed hebben op de stofwisseling van de monoaminen. Na toediening van MAO-remmers bij proefdieren is dan ook gebleken dat de concentratie aan monoaminen in het centrale zenuwstelsel sterk stijgt; verscheidene honderden MAO-remmers zijn in de laatste jaren door farmaceutische laboratoria gesynthetiseerd, een klein percentage ervan is van blijvende waarde gebleken. De meeste thans in gebruik zijnde MAO-remmers zijn hydrazinederivaten die worden gekenmerkt door de hydrazinegroep.Ze bezitten zoals de naam aangeeft, het vermogen het enzym MAO te blokkeren. Er bestaat een duidelijk antagonisme tussen de MAO-remmers en reserpine wat betreft het vermogen monoaminen vrij te maken; de MAO-remmers zijn echter niet in staat de vrijmaking van monoaminen door sympathicomimetica te blokkeren.
Tenminste twee mechanismen spelen een rol bij de opname van de monoaminen in de sympathische neuronen. Eén systeem zou verantwoordelijk zijn voor het transport door het membraan van het neuron; een ander voor de opname in de granulae. Het laatste mechanisme zou nu geblokkeerd worden door reserpine, waardoor een extra hoeveelheid monoaminen zich ophoogt in het cytoplasma, hetgeen leidt tot een toename van de oxydatieve desaminering en een verhoogde afgifte van de afvalproducten aan de extracellulaire ruimte.
COMT
Catechol-O-methyl-transferase, het enzym dat de catecholaminen inactiveert buiten de zenuwen, komt overal in het lichaam en ook in de hersenen voor, maar de functie ervan is verschillend van MAO. Ook de activiteit van COMT kan geremd worden en het farmacon pyrogallol is in dit opzicht actief. (122)
Overzicht van de werking van de catecholaminen
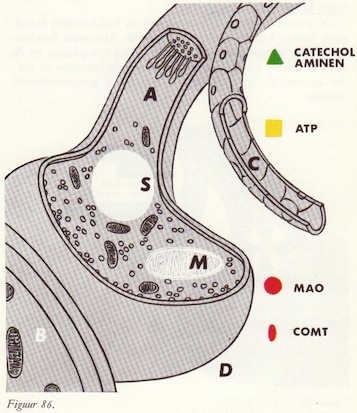 Teneinde een duidelijk beeld te krijgen van het werkingsmechanisme van de monoaminergische transmitters en hun mogelijk klinische betekenis, zal in een aantal figuren een schematisch overzicht gegeven worden van de synthese en productie, de opslag en afgifte, en de remmers van deze mechanismen. Figuur 86 bevat alle structurele elementen waarin en waartussen de verschillende stappen van synthese en afbraak zich afspelen.
Teneinde een duidelijk beeld te krijgen van het werkingsmechanisme van de monoaminergische transmitters en hun mogelijk klinische betekenis, zal in een aantal figuren een schematisch overzicht gegeven worden van de synthese en productie, de opslag en afgifte, en de remmers van deze mechanismen. Figuur 86 bevat alle structurele elementen waarin en waartussen de verschillende stappen van synthese en afbraak zich afspelen.A. stelt zowel een postganglionair sympathisch neuron als een schakelcel in een van de vele circuits in het centrale zenuwstelsel voor, die aan het uiteinde een van de monoaminergische transmitters afscheidt. Mitochondria en synaptische blaasjes of granulae spelen de belangrijkste rol bij de transmitteroverdracht; een mitochondrium (M) en een synaptisch blaasje (S) zijn vergroot weergegeven.
B. is een aanduiding van de effector (kliercel of spiervezel, of elk ander weefselelement waarop de monoaminergische transmitters hun invloed kunnen uitoefenen). Ook kan B het postsynaptische element van een neuronenketen voorstellen.
C. is een capillair waarin de basiselementen voor de transmitters of de complete transmittermoleculen worden aangevoerd.
D. is de extracellulaire ruimte en tevens wordt hierin het voorkomen van de afvalproducten in de urine aangegeven.
De catecholaminen kunnen zowel endogeen (in het neuron zelf) geproduceerd worden als exogeen (uit de circulatie) aangevoerd worden. Noradrenaline, de belangrijkste catecholamine in het perifere sympathische zenuwstelsel, wordt in alle delen van het neuron gevonden. De concentratie is laag in het cellichaam (10-500 μg/g) maar zeer hoog in de uiteinden van het neuron (plm. 10.000 μg/g). De uitstulpingen van het postganglionaire neuron, die op vele plaatsen in de autonome plexus van de organen worden gezien, bevatten eveneens een hoge concentratie catecholaminen. (124)
Catecholaminendepots
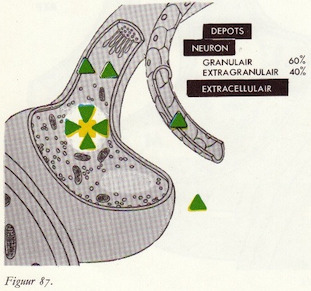 Minutieus biochemisch en elektronenmicroscopisch onderzoek heeft aangetoond, dat er tenminste drie depots bestaan waarin catecholaminen voorkomen: in het neuron een granulair depot en een cytoplasmatisch of extragranulair depot, en buiten het neuron een extracellulair depot. Van de neuronale catecholaminen bevindt zich ongeveer 40% in het extragranulaire of 'vrije' depot en 60% in het granulaire of 'gebonden' depot. De op deze wijze gebonden transmitters zijn fysiologisch niet actief maar potentieel wel. Een klein deel van de 60% kan gemakkelijk vrijgemaakt worden, het grootste deel is echter gebonden aan ATP-moleculen in een verhouding van 4:1.
Minutieus biochemisch en elektronenmicroscopisch onderzoek heeft aangetoond, dat er tenminste drie depots bestaan waarin catecholaminen voorkomen: in het neuron een granulair depot en een cytoplasmatisch of extragranulair depot, en buiten het neuron een extracellulair depot. Van de neuronale catecholaminen bevindt zich ongeveer 40% in het extragranulaire of 'vrije' depot en 60% in het granulaire of 'gebonden' depot. De op deze wijze gebonden transmitters zijn fysiologisch niet actief maar potentieel wel. Een klein deel van de 60% kan gemakkelijk vrijgemaakt worden, het grootste deel is echter gebonden aan ATP-moleculen in een verhouding van 4:1.De granulae zijn van fysiologisch en biochemisch standpunt bezien bijzonder belangwekkend. Allereerst beschermen ze de neurotransmitters tegen in het cytoplasma en de mitochondriën voorkomende destruerende enzymen en verder kunnen de transmitters in hoge mate geconeentreerd worden. (126)
Opname van catecholaminen in het neuron
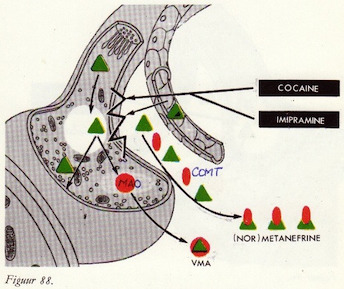 De neuronen bezitten een zeer effectief mechanisme voor het opnemen van catecholaminen uit de bloedbaan en de extracellulaire ruimte. Hierbij spelen zgn. metabole pompen, die in het neuronale membraan gelokaliseerd zijn, een essentiële rol. Een soortgelijk pompmechanisme zijn we reeds tegengekomen bij het handhaven van het natrium- en kaliumevenwicht tijdens de rustpotentiaal van het neuron. Het pompsysteem van het neuron kan geblokkeerd worden door o.a. cocaine en imipramine. Wanneer dit het geval is, wordt de extracellulaire noradrenaline afgebroken door COMT, waardoor in de urine een toename wordt gevonden van normetanefrine en metanefrine.
De neuronen bezitten een zeer effectief mechanisme voor het opnemen van catecholaminen uit de bloedbaan en de extracellulaire ruimte. Hierbij spelen zgn. metabole pompen, die in het neuronale membraan gelokaliseerd zijn, een essentiële rol. Een soortgelijk pompmechanisme zijn we reeds tegengekomen bij het handhaven van het natrium- en kaliumevenwicht tijdens de rustpotentiaal van het neuron. Het pompsysteem van het neuron kan geblokkeerd worden door o.a. cocaine en imipramine. Wanneer dit het geval is, wordt de extracellulaire noradrenaline afgebroken door COMT, waardoor in de urine een toename wordt gevonden van normetanefrine en metanefrine.Is de noradrenaline of een van de andere monoaminen eenmaal door het neuronale membraan getransporteerd, dan wordt het opgenomen in het extragranulaire of het granulaire depot. Hierbij spelen een groot aantal factoren een rol, zoals actieve opname, passieve diffusie, etc. (127)
Opname van catecholaminen in de granulae
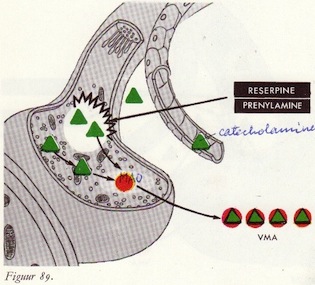 Ook bij het transport van de catecholaminen van het extragranulaire depot naar de granulae spelen pompmechanismen een rol. Ook deze pomp is een enzymatisch systeem en de functie is afhankelijk van ATP en Mg. Het granulaire opname mechanisme wordt geremd door reserpine, tot voor kort de meest effectieve remmer. Een nieuwe remmer, prenylamine, werkt effectiever en blokkeert niet alleen de opname van de catecholaminen in de granulae, maar veroorzaakt tevens een depletie van de aanwezige catecholaminen. De in het cytoplasma opgehoopte neurotransmitters worden nu door de mitochondriën opgenomen en door MAO afgebroken. De noradrenergische invloed op de effector wordt zodoende voor een groot deel uitgeschakeld. (128)
Ook bij het transport van de catecholaminen van het extragranulaire depot naar de granulae spelen pompmechanismen een rol. Ook deze pomp is een enzymatisch systeem en de functie is afhankelijk van ATP en Mg. Het granulaire opname mechanisme wordt geremd door reserpine, tot voor kort de meest effectieve remmer. Een nieuwe remmer, prenylamine, werkt effectiever en blokkeert niet alleen de opname van de catecholaminen in de granulae, maar veroorzaakt tevens een depletie van de aanwezige catecholaminen. De in het cytoplasma opgehoopte neurotransmitters worden nu door de mitochondriën opgenomen en door MAO afgebroken. De noradrenergische invloed op de effector wordt zodoende voor een groot deel uitgeschakeld. (128)De afgifte van noradrenaline
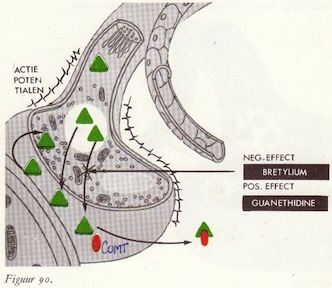 De noradrenaline in de postganglionaire sympathische neuronen wordt onder normale omstandigheden vrijgemaakt door de actiepotentialen, die in het neuron opgewekt worden. Die actiepotentialen zijn in staat vooral de granulae die dicht bij het membraan gelegen zijn, geheel of gedeeltelijk te ledigen. Er vindt geen afgifte plaats uit het extragranulaire (zgn. vrije) depot. Dit fenomeen vertoont dus grote overeenkomst met het vrijkomen van acetylcholine uit de cholinergische neuronen en de neuromusculaire synaps.
De noradrenaline in de postganglionaire sympathische neuronen wordt onder normale omstandigheden vrijgemaakt door de actiepotentialen, die in het neuron opgewekt worden. Die actiepotentialen zijn in staat vooral de granulae die dicht bij het membraan gelegen zijn, geheel of gedeeltelijk te ledigen. Er vindt geen afgifte plaats uit het extragranulaire (zgn. vrije) depot. Dit fenomeen vertoont dus grote overeenkomst met het vrijkomen van acetylcholine uit de cholinergische neuronen en de neuromusculaire synaps.De afgifte van catecholaminen door actiepotentialen kan nagebootst worden door guanethidine en het mechanisme wordt geblokkeerd door o.a. bretylium. Ca-ionen en waarschijnlijk acetylcholine zijn noodzakelijk voor de afgifte uit het granulaire depot.
Sympathicomimetica hebben een analoog effect als de actiepotentialen. Een groot percentage van het vrijgekomen noradrenaline wordt weer opgenomen door de zenuw. Kleine hoeveelheden noradrenaline worden voortdurend spontaan vrijgemaakt. (129)
MAO en MAO-remmers
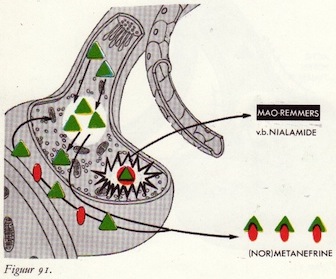 Het mitochondriale MAO inactiveert alleen de cytoplasmatische catecholaminen. Na binnenkomst in de intraneuronale ruimte staan er twee wegen open: opname in het granulaire depot of inactivering door MAO. Na desaminering door MAO worden de geïnactiveerde producten door het neuron afgegeven en neemt het gehalte van VMA in de extracellulaire ruimte toe.
Het mitochondriale MAO inactiveert alleen de cytoplasmatische catecholaminen. Na binnenkomst in de intraneuronale ruimte staan er twee wegen open: opname in het granulaire depot of inactivering door MAO. Na desaminering door MAO worden de geïnactiveerde producten door het neuron afgegeven en neemt het gehalte van VMA in de extracellulaire ruimte toe.MAO-remmers verhinderen de inactivering van de catecholaminen door MAO, waardoor een toename van zowel het extragranulaire als het granulaire depot optreedt. Daar de noradrenaline niet meer door MAO geïnactiveerd wordt, is een afname van VMA het gevolg; de COMT blijft echter beschikbaar, waardoor normetanefrine toeneemt.
De MAO-remmers vinden vooral toepassing in de neuropsychiatrische kliniek bij de behandeling van vitale depressies. De vraag naar een eventuele samenhang tussen stofwisselingsprocessen van de monoaminen enerzijds en de invloed van deze en soortgelijke neurotransmitters op de psyche anderzijds, wordt in vele laboratoria onderzocht. (130)
Reserpine-effect
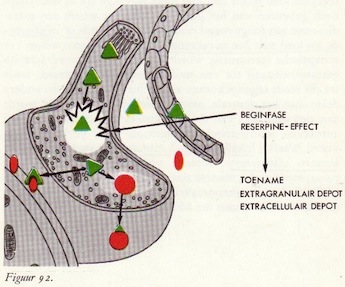 Het effect van reserpine op de stapeling en afgifte van catecholaminen uit de diverse depots is een van de belangrijkste grondslagen gebleken voor het begrijpen van het aangrijpingspunt van een aantal geneesmiddelen (o.a. prenylamine) die op de een of andere manier de monoaminen beïnvloeden. Het centrale effect van prenylamine is vergelijkbaar met reserpine.
Het effect van reserpine op de stapeling en afgifte van catecholaminen uit de diverse depots is een van de belangrijkste grondslagen gebleken voor het begrijpen van het aangrijpingspunt van een aantal geneesmiddelen (o.a. prenylamine) die op de een of andere manier de monoaminen beïnvloeden. Het centrale effect van prenylamine is vergelijkbaar met reserpine.In het beginstadium van het reserpine-effect treedt een blokkade op van het transport van het cytoplasmatische naar het granulaire depot. De inactivering van de catecholaminen door MAO neemt toe, evenals de afgifte aan de extracellulaire ruimte. Actiepotentialen hebben nog wel een afgifte van granulaire catecholaminen tengevolge, daar nog steeds een voorraad in de granulae aanwezig is.
Na enige tijd zijn de depots in de granulae uitgeput, waardoor de actiepotentialen geen afgifte van noradrenaline meer veroorzaken. Het postganglionaire sympathische neuron is dan niet of nauwelijks meer in staat zijn noradrenergische invloed op de effectoren uit te oefenen. (131)
De organisatie van het autonome zenuwstelsel
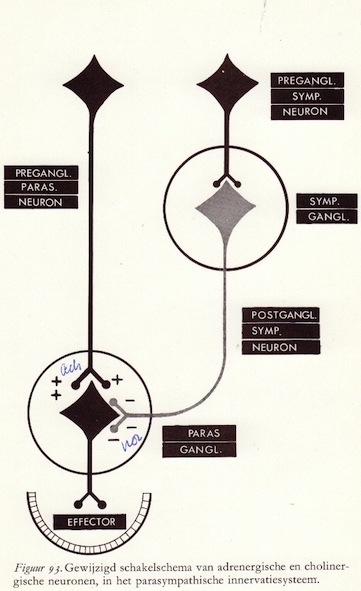 Met zeer gevoelige fluorescentietechnieken is men in staat geweest neerslagen van catecholaminen in histologische preparaten aan te tonen op plaatsen waar dit tot nu toe niet werd vermoed en zelfs zeer onwaarschijnlijk werd geacht. Verschillende kerngebieden in het centrale zenuwstelsel bleken rijk aan catecholaminen te zijn; maar ook in het perifere autonome zenuwstelsel werden enkele verrassende ontdekkingen gedaan. Zo kwam onomstotelijk de aanwezigheid vast te staan van sympathische postganglionaire eindigingen in parasympathische perifere ganglia: de cellichamen synapteren met cholinerge én noradrenerge axonen. Het postganglionaire parasympathische neuron staat dus niet alleen onder controle van het acetylcholine afscheidende preganglionaire parasympathische neuron, maar tevens van het noradrenaline afscheidende postganglionaire sympathische neuron.
Met zeer gevoelige fluorescentietechnieken is men in staat geweest neerslagen van catecholaminen in histologische preparaten aan te tonen op plaatsen waar dit tot nu toe niet werd vermoed en zelfs zeer onwaarschijnlijk werd geacht. Verschillende kerngebieden in het centrale zenuwstelsel bleken rijk aan catecholaminen te zijn; maar ook in het perifere autonome zenuwstelsel werden enkele verrassende ontdekkingen gedaan. Zo kwam onomstotelijk de aanwezigheid vast te staan van sympathische postganglionaire eindigingen in parasympathische perifere ganglia: de cellichamen synapteren met cholinerge én noradrenerge axonen. Het postganglionaire parasympathische neuron staat dus niet alleen onder controle van het acetylcholine afscheidende preganglionaire parasympathische neuron, maar tevens van het noradrenaline afscheidende postganglionaire sympathische neuron.Dit heeft uiteraard belangrijke consequenties voor de organisatie en de integratiecapaciteit van het perifere parasympathische neuron. De exciterende activiteit van het acetylcholine speelt dan niet meer de enige rol in het synaptische gebeuren van het ganglion, maar er wordt een extra eigenschap aan toegevoegd door de modulerende of inhiberende invloed van noradrenaline. Van een eenvoudige 1:1 synaptische transmissie wordt de informatie-overdracht nu getransformeerd tot een multifunctioneel systeem, zoals we dit reeds zijn tegengekomen in het ruggemerg en andere delen van het centrale zenuwstelsel.
Mogelijkerwijs wordt dan ook een deel van de integratieve capaciteiten van het autonome zenuwstelsel niet centraal maar perifeer uitgevoerd, waarbij lokale feedback-circuits een sturende functie kunnen uitoefenen. Depletie van catecholaminen in het parasympathische neuron zou verder een parasympathische facilitatie kunnen veroorzaken, die buiten de controle van de centrale mechanismen om loopt. (132)
terug naar de Inhoud
5. Lijst van termen
Teneinde het lezen van de tekst te vergemakkelijken wordt hieronder een verklaring van de 50 meest gebruikte termen gegeven.
- Acetylcholine. Biochemische stof die aan de uiteinden van bepaalde zenuwen vrijkomt. Speelt een belangrijke rol bij de overdracht van impulsen tussen zenuwcellen en tussen zenuwcellen en dwarsgestreepte spieren. Wordt afgebroken door het enzym choline-esterase.
- Actiepotentiaal. Potentiaalverschil tussen het actieve en niet-actieve deel van een zenuwcel of spiervezel.
- Actine. Een van de contractiele eiwitten in het spierplasma.
- ADP (adenosinedifosfaat). Biochemische stof die een belangrijke rol speelt bij energie-overdracht, zoals bij spiercontractie. Wordt gevormd door hydrolyse van ATP (zie aldaar) met behulp van het enzym adenosinetransfosfatase.
- Adrenaline (epinefrine). a. een hormoon, afkomstig uit het bijniermerg; b. een neurotransmitter, die aan de uiteinden van sympathische postganglionaire zenuwen wordt vrijgemaakt, maar ook aan de uiteinden van verschillende neuronen in het centrale zenuwstelsel.
- Afferent (aanvoerend). Afferente zenuwen (-vezels) vervoeren prikkels van een receptor naar het centrale zenuwstelsel.
- Aminen (amino-basen). Worden van ammoniak afgeleid door 1, 2 of 3 waterstofatomen te vervangen door alkylen (koolwaterstof-resten).
- Amplitudo. Hoogte, uitslag (van het meetapparaat), afstand tussen de uiterste punten van een golfbeweging, loodrecht op de bewegingsrichting.
- Antagonistisch (elkaar tegenwerkend). In de spierfysiologie worden onder antagonisten verstaan spieren, waarvan de werking aan elkaar tegengesteld is.
- ATP (adenosinetrifosfaat). Biochemische verbinding die een belangrijke rol speelt bij de celstofwisseling. ATP bezit twee energierijke fosforzuur-bindingen; de spier krijgt zijn energie uit de hydrolyse (ontleding van organische moleculen onder afgifte van water) van ATP in ADP (zie aldaar). Neuronale membranen bevatten een hoge concentratie ATP. (137)
- Axon (ook: neuriet). De langste en in cytologische karakterestieken van de rest van het neuron afwijkende uitloper van een zenuwcel. De prikkelgeleiding is in de meeste gevallen en onder normale omstandigheden van de zenuwcel af. Een bundel axonen vormt een zenuw.
- Cholesterol. Een lipoïd dat voorkomt in vele organen, bijvoorbeeld bloed, zenuwen, lever, nieren etc. (lipoïden: vetachtige bestanddelen die echter niet, zoals vetten, kunnen worden verzeept maar wel in kokende alcohol en ether oplosbaar zijn).
- Cholinerg. Zenuwvezels, die als ze geactiveerd worden acetylcholine afscheiden, worden cholinerg genoemd.
- Cortex. Schors, zoals cortex cerebri: schors van de grote hersenen, en cortex cerebelli: schors van de kleine hersenen.
- Dendriet. In vergelijking met het axon meestal korte uitloper van het zenuwcellichaam, cytologisch met dat cellichaam veel punten van overeenkomst vertonend. De geleiding is naar de zenuwcel toe. Het aantal per neuron varieert van 1 tot 12.
- Depolarisatie. Een in de neurofysiologie gebruikte term die duidt op een toestand waarbij de membraanpotentiaal minder negatief wordt.
- Effector. Een spier of klier die, na ontvangst van een impuls van de bijbehorende afferente zenuw, een toestandsverandering in zijn omgeving teweegbrengt.
- Efferent (ook: wegvoerend, centrifugaal). Efferente zenuwen (-vezels) voeren prikkels van het centrale zenuwstelsel naar de periferie. Zeer veel efferente zenuwvezels zijn motorisch.
- Elektroencefalogram (EEG). De kromme die men verkrijgt bij het registreren van de elektrische verschijnselen in de hersenen.
Het alfa-ritme (845 perioden per sec.) is het meest voorkomende EEG. De veranderingen in het EEG stellen ons in staat bepaalde ziekten te diagnosticeren en beschadigingen van de hersenen te lokaliseren.
- Enzym. Eiwit met een werkzaam gedeelte (co-enzym) dat in staat is een biochemische reactie te katalyseren (versnellen).
- EPSP. Exciterende postsynaptische potentiaal; een actieve elektrische verandering van een synaptisch membraan van de postsynaptische cel, die door een exciterende presynaptisehe impuls wordt opgewekt. De postsynaptische cel wordt gedepolariseerd. (138)
- Ganglion. Opeenhoping van zenuwcellichamen buiten het centrale zenuwstelsel (binnen het centrale zenuwstelsel: nucleus of kern).
- Generatorpotentiaal. Kleine depolariserende potentiaalverschillen, opgewekt in de receptorcellen.
- Gliacel. Ondersteunend en voedend element van het niet-neuronale weefsel van het zenuwstelsel.
Hyperpolarisatie. Een in de neurofysiologie gebruikte term die duidt op een toestand, waarbij de membraanpotentiaal meer negatief wordt.
- Impuls (zenuw-). Signaal dat door zenuwcellen en hun uitlopers wordt voortgeleid. De passage van een impuls langs de zenuwcel blijkt uit een plotselinge verandering van het elektrische en het ionenevenwicht dat tijdens rust (rustpotentiaal) bestaat tussen het inwendige en uitwendige van de zenuwcel.
- Inhibitie (remming). Onderdrukking van een proces of verschijnsel.
- Ion. Atoom of molecuul dat een of meer elektronen heeft opgenomen of afgegeven; het heeft hierdoor een elektrische lading (positief of negatief) gekregen.
- IPSP. Inhiberende postsynaptische potentiaal: een actieve elektrische verandering van een synaptisch membraan van de postsynaptische cel, die door een inhiberende presynaptische impuls wordt opgewekt. De postsynaptische cel wordt gehyperpolariseerd.
- Membraan. Vliesachtige structuur die het cytoplasma van iedere dierlijke of plantaardige cel omgeeft. Dit membraan is selectief permeabel en speelt een belangrijke rol bij de celstofwisseling.
- Mitochondria. Langwerpige of ronde lichaampjes in het cytoplasma die men bij alle cellen heeft kunnen aantonen, behalve bij bacteriën. Ze bevatten bijna alle enzymen voor de celademhaling; ze vormen dus de energiecentrales (ATP-aanmaak) voor de gehele cel.
- Motorische voorhoorncel. Zenuwcel, in de voorhoorn van het ruggemerg gelegen. Het axon van deze cel verzorgt de motorische innervatie van de dwarsgestreepte spieren.
- Myeline. Stof in de myelineschede van het axon, samengesteld uit vetten en eiwitten.
- Myosine. Een van de contractiele eiwitten in het spierplasma.
- Neuron. Een zenuwcel met zijn uitlopers. Het neuron vormt de bouwsteen van het zenuwstelsel. (139)
- Neurotransmitter. Biochemische stof, verantwoordelijk voor het overbrengen van de impuls van de ene zenuwcel op de andere.
- Noradrenaline (norepinefrine). In tegenstelling tot adrenaline draagt het stikstof geen -CH3 groep maar slechts een -H atoom). a. een hormoon, ook afkomstig uit het bijniermerg, maar in veel mindere mate dan adrenaline; b. een neurotransmitter die, evenals adrenaline, aan de uiteinden van sympathische postganglionaire zenuwen wordt vrijgemaakt, maar ook aan de uiteinden van verschillende neuronen in het centrale zenuwstelsel.
- Nucleïnezuren. Moleculen die voor het leven onmisbaar zijn, bouwstenen van het DNA. Het zijn eiwitten, opgebouwd uit een keten bestaande uit een zeer groot aantal schakels: de nucleotiden. Er zijn twee soorten nucleïnezuren: het desoxyribonucleïnezuur (DNA) en het ribonucleïnezuur (RNA). Het DNA komt alleen in de kern voor, het is het voornaamste bestanddeel van de chromosomen; het verleent aan de cel de erfelijke eigenschappen. Het RNA bevindt zich zowel in de kern als in het cytoplasma; het speelt een belangrijke rol bij de eiwitsynthese.
- Oragnellen. Zelfstandige kleine orgaantjes in een cel.
- Permeabiliteit. Het vermogen van stoffen om een celmembraan te passeren. Het celmembraan laat slechts die stoffen door die nuttig zijn voor het functioneren van de cel.
- Postganglionair. Van de twee achter elkaar geschakelde neuronen die het perifere deel van het autonome zenuwstelsel vormen, heet die waarvan de oorsprongcel in een perifeer ganglion ligt, postganglionair.
- Preganglionair. Van de twee achter elkaar geschakelde neuronen die het perifere deel van het autonome zenuwstelsel vormen, heet die waarvan de oorsprongcel in het centrale zenuwstelsel ligt, preganglionair.
- Receptor. Molecuul of orgaan, gespecialiseerd in het ontvangen van informatie uit de buiten- en binnenwereld. Wordt ook wel in engere zin gebruikt m.b.t. die cellen van de receptor, die de genoemde informatie 'vertalen' en doorgeven aan de sensibele zenuwen.
- Refractaire periode, absolute - : de periode die volgt op de excitatie van een zenuwcel gedurende welke geen prikkel, hoe sterk ook, een volgende actiepotentiaal kan opwekken; relatieve - : de periode die volgt op de excitatie van een zenuwcel, waarbij een sterkere prikkel wel een actiepotentiaal kan opwekken. (140)
- Ribonucleïnezuren. Zie nucleïnezuren.
- Sarcoplasma. Niet gedifferentieerd protoplasma der spiervezels, tussen de fibrillen en rondom de kernen.
- Servomechanisme. Een apparaat dat werkt op basis van verkregen informatie, met als doel de informatie die in de toekomst zal worden verkregen in overeenstemming met een bepaalde opdracht te veranderen; een thermostaat, die de temperatuur van de kamer meet en de centrale verwarming zo regelt dat de temperatuur constant op een bepaalde waarde blijft, is een servomechanisme.
- Synaps. Plaats waar twee zenuwcellen met elkaar in functioneel contact treden.
- Terugkoppeling (negatieve). Het aanwenden van de output van een systeem om de input van dat systeem zo te veranderen, dat het verschil tussen de input en de voorgeschreven input wordt verkleind (tegenkoppeling). (141)
terug naar het literatuuroverzicht
^